Plk1 coordonne l’entrée et la progression mitotiques en amplifiant le signal cycline B/CDK1, en organisant la bipolarité et la dynamique du fuseau, puis en préparant la zone médiane et la cytokinèse.
Vue d'ensemble de Plk1
Plk1 (Polo-like Kinase 1) est une kinase sérine/thréonine centrale qui n’initie pas l’entrée en mitose mais en assure la coordination spatio-temporelle, depuis la transition G2/M jusqu’à la préparation de la cytokinèse.
1. Son rôle principal est de relayer, amplifier et spatialiser le signal mitotique porté par le complexe cycline B/CDK1 (MPF), en l’articulant avec l’organisation cellulaire :
Plk1 se distingue ainsi des kinases déclencheuses par une action séquentielle et localisée lui conférant un rôle d’intégrateur enzymatique du réseau mitotique plutôt que de régulateur autonome du cycle cellulaire.
3. Le tableau suivant, qui n’a pas vocation à être exhaustif, montre les principaux substrats phosphorylés par Plk1 au cours du cycle mitotique, classés selon la phase mitotique dominante, la localisation subcellulaire et la conséquence fonctionnelle principale, afin de mettre en évidence son rôle transversal.
2. L’activité du domaine kinase reste cependant insuffisante en l’absence d’un ciblage spatial approprié, soulignant la dépendance fonctionnelle de Plk1 à son domaine Polo-Box C-terminal pour la reconnaissance des substrats et sa localisation subcellulaire.
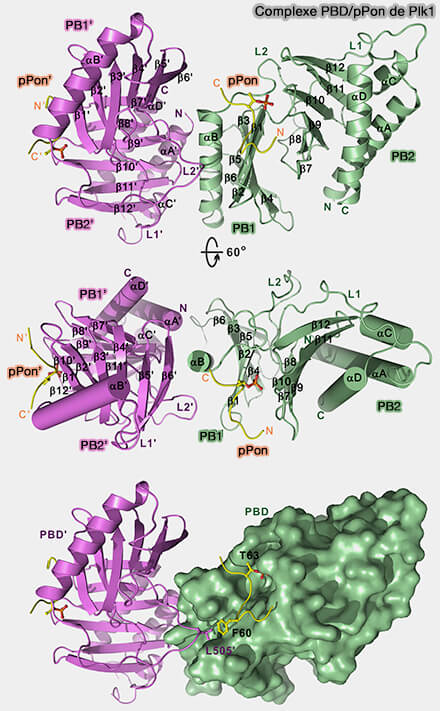
Complexe PBD/pPon de Plk1
(Figure : vetopsy.fr d'après Zhu et coll)
PB1 contient la poche principale de liaison aux résidus phospho-sérine ou phospho-thréonine présents sur les protéines cibles ou adaptatrices.
PB2, bien qu'apparenté, joue un rôle architectural et modulateur, contribuant à la stabilité du complexe et à la spécificité contextuelle du ciblage.
Remarque : sur la figure, pPon est un peptide phosphorylé utilisé comme modèle pour l’étude de la reconnaissance phospho-dépendante par le domaine Polo-Box.
Les assemblages en dimères de dimères observés en cristallographie reflètent des contacts de cristallisation et non une oligomérisation stable in vivo.
La charge positive partielle de la gorge PB1-PB2 favorise la stabilisation du groupement phosphate et impose une orientation précise du peptide, conditionnant la fidélité de reconnaissance.
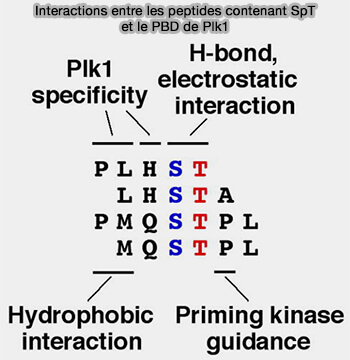
Interactions peptides à SpT et PBD de Plk1
(Figure : vetopsy.fr d'après Yun et coll) Cette poche n’apparaît que dans des conformations ouvertes du PBD et module la sélectivité de liaison de Plk1 envers différents partenaires.
Ce double mode de liaison, gorge + poche hydrophobe, permet à Plk1 de discriminer entre différents sous-ensembles de substrats.
3. Ce mode de reconnaissance confère à Plk1 une capacité de ciblage séquentiel vers différentes plateformes subcellulaires, lui permettant de se localiser successivement :
4. Ainsi, le domaine Polo-Box confère à Plk1 un mode d’action contextuel en assurant un adressage spatio-temporel phospho-dépendant qui restreint l’activité kinase aux plateformes cellulaires définies par l’état mitotique local et l’architecture cellulaire, plutôt que par une régulation autonome de type transcriptionnel ou cyclique.
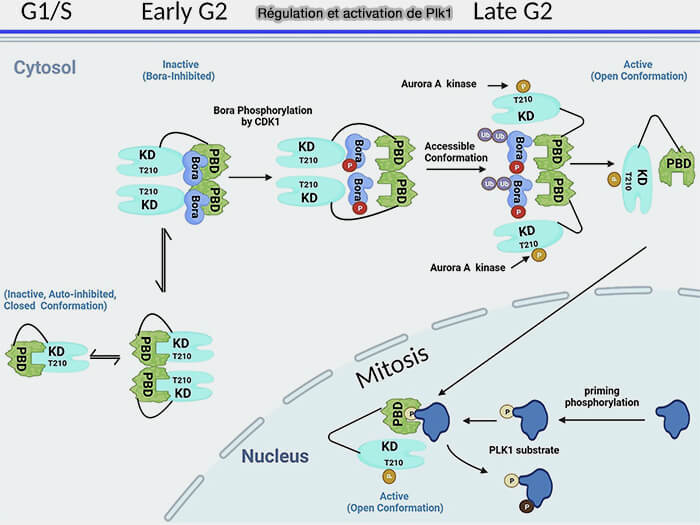
1. En interphase et jusqu'à la fin de la phase S, Plk1 est maintenue dans une conformation autoinhibée grâce à des interactions intramoléculaires entre son domaine Polo-Box (PBD) et son domaine kinase, ainsi que par l’existence de formes dimériques fonctionnellement inhibées.
2. En phase G2 précoce, la protéine Bora, exprimée et accumulée spécifiquement pendant cette phase, agit comme cofacteur d’activation en se liant à Plk1 et en stabilisant un état pré-mitotique réversible, qui limite toute activité kinase prématurée tout en préparant la protéine à une activation rapide.
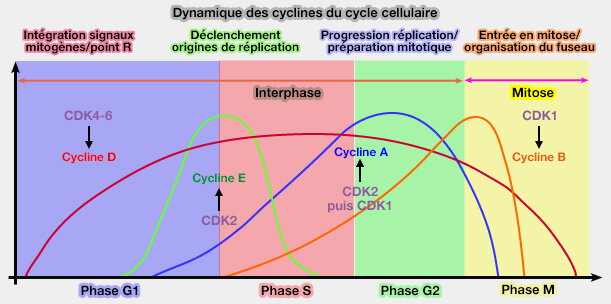
Dynamique des cyclines du cycle cellulaire
(Figure : vetopsy.fr)
Cette phosphorylation stabilise la protéine et favorise la formation d’un complexe transitoire Plk1-Bora, créant un état conformationnel permissif à la phosphorylation du site d’activation Thr210 par Aurora A.
Tant que Bora n’est pas phosphorylée par CDK1, elle ne favorise pas efficacement l’activation de Plk1.
une transition conformationnelle majeure, rendant la boucle d’activation catalytiquement compétente,
la dissociation des formes oligomériques pour une forme monomérique fonctionnelle.
a. Cette phosphorylation est stable dans le contexte mitotique et n’est inversée qu’en sortie de mitose par l’action des phosphatases PP1 et PP2A.
b. Parallèlement, la poursuite de la phosphorylation de Bora sur plusieurs sites, par CDK1 et probablement d’autres kinases mitotiques, génère un phospho-dégron reconnu par le complexe SCFβ-TrCP, conduisant à sa polyubiquitination et à sa dégradation protéasomale.
La dégradation de Bora met fin à son rôle de cofacteur d’activation initial sans affecter l’état actif de Plk1, désormais maintenu dans le contexte mitotique.
Régulation et activation de Plk1 (Polo-like Kinase 1)
(Figure : vetopsy.fr d'après Chapagai et coll)
Remarque : dans la figure ci-dessus, la présence d’une enveloppe dans un schéma associé à une activité nucléaire de Plk1 doit être comprise comme une simplification graphique, et non comme une affirmation d’un passage actif de Plk1 à travers des pores nucléaires pendant la prométaphase.
a. Une fois phosphorylée sur Thr210, Plk1 est recrutée, via son domaine Polo-Box, sur des plateformes subcellulaires pré-phosphorylées où elle phosphoryle localement des régulateurs du fuseau, des attachements kinétochoriens et de la zone médiane.
b. Parallèlement, Plk1 phosphoryle Nlp (Ninein-like protein), entraînant sa dissociation du centrosome. et levant ainsi un frein structurel qui contribue à l’expansion du PCM et favorise la nucléation mitotique massive des microtubules.
Cette maturation centrosomale prépare la formation d’asters mitotiques robustes, indispensables à la migration et à la séparation ultérieure des pôles.
2. Parallèlement, Plk1 amplifie la séparation des centrosomes initiée par Nek2 en activant les forces mécaniques d’écartement.
Ainsi, en interphase tardive, Plk1 ne construit pas encore le fuseau mitotique, mais établit un état cytoplasmique et centrosomal permissif, caractérisé par des pôles compétents, une nucléation microtubulaire accrue et des forces d’écartement prêtes à être mobilisées lors de l’engagement mitotique.
1. Plk1 renforce l’engagement irréversible en mitose en contribuant à l’amplification et à la propagation spatiale de l’état mitotique depuis les centrosomes vers le reste de la cellule.
Elle joue un rôle clé dans le renforcement des boucles de rétroactivation du réseau mitotique, en particulier l’axe MPF ➞ Cdc25 ➞ MPF, accélérant la montée globale de l’activité CDK1.
En parallèle, elle participe à la levée progressive des freins à l’entrée en mitose, notamment par l’inhibition fonctionnelle de Wee1 et Myt1 (PKMYT1), supprimant les phosphorylations inhibitrices résiduelles de CDK1 et favorisant l’engagement irréversible de la cellule dans l’état mitotique.
2. Sur le plan organisationnel, Plk1 ne prépare plus les pôles mais déclenche la mise en œuvre effective de la mise en place du fuseau mitotique naissant en orchestrant plusieurs processus clés :
l’organisation rapide et coordonnée des microtubules polaires, par la phosphorylation de protéines associées aux microtubules et aux pôles, telles que Nde1/Ndel1, TACC3 et CLIP-170, permettant l’établissement d’une architecture bipolaire dynamique,
la synchronisation des compartiments nucléaire et cytoplasmique, par la phosphorylation de régulateurs du cycle cellulaire, en particulier Cdc25, via l’activation de Cdc25 et l’inhibition fonctionnelle de Wee1 et Myt1, assurant une montée cohérente et globale de l’état mitotique.
stimulant la dynamique des microtubules, notamment par la phosphorylation de protéines associées aux pôles et aux microtubules telles que Nde1/Ndel1, TACC3 et CLIP-170, ce qui accroît l’instabilité contrôlée des extrémités et facilite l’exploration du volume cellulaire.
soutenant la maturation fonctionnelle des pôles centrosomaux par son action sur des composants du matériel péricentriolaire, renforçant la nucléation et l’organisation des microtubules polaires.
b. Cette coordination favorise l’établissement et le maintien d’une architecture bipolaire du fuseau, condition indispensable à la capture, au repositionnement et au réalignement progressif des chromosomes.
2. Plk1 assure la cohérence temporelle de l’état mitotique en prométaphase en contribuant au maintien et à la propagation du signal mitotique imposé par le complexe cycline B/CDK1 (MPF), empêchant toute sortie prématurée de mitose tant que les attachements kinétochoriens ne sont pas stabilisés.
Plk1 et métaphase
1. En métaphase, Plk1 contribue au maintien fonctionnel de l’état métaphasique et à la robustesse du fuseau.
Plk1 contribue à l’établissement d’un contexte permissif à la sortie de métaphase en préparant fonctionnellement les complexes impliqués dans la transition métaphase-anaphase, sans encore déclencher l’activation de l’APC/CCdc20.
2. Plk1 agit ainsi comme une kinase de consolidation, assurant la fiabilité mécanique du fuseau et la cohérence enzymatique de l’état métaphasique, tout en maintenant la cellule prête à engager rapidement la transition vers l’anaphase lorsque les conditions sont réunies.
Plk1 en anaphase et télophase
1. En anaphase et en télophase, Plk1 n’initie plus les transitions mitotiques, mais accompagne les phases tardives de la division en coordonnant le remodelage du fuseau et la préparation spatiale de la cytokinèse.
Plk1 participe à la réorganisation du fuseau central et à la définition de la zone médiane par la phosphorylation de composants clés du complexe centralspindlin et de ses régulateurs.
Elle module notamment l’activité et la localisation de PRC1 et de MKLP1 (KIF23), favorisant l’alignement, la stabilisation contrôlée et la cohérence des faisceaux de microtubules antiparallèles.
En parallèle, Plk1 ajuste les propriétés mécaniques du fuseau tardif par la phosphorylation de protéines associées aux microtubules polaires et interpolaires, telles que KIF4A, CLASP1/2 et TACC3, contribuant au contrôle de la longueur des microtubules, à leur stabilité et à la coordination avec l’allongement du fuseau en anaphase B.
2. Cette activité s’accompagne d’une relocalisation progressive de Plk1 vers la zone médiane, où elle contribue à :
préparer l’assemblage du site de cytokinèse avant la formation du corps intermédiaire.
L’extinction progressive de l’activité de Plk1 ne constitue pas un mécanisme autonome, mais s’inscrit dans la bascule globale kinases-phosphatases qui gouverne la sortie de mitose, conduisant au démantèlement du fuseau tardif, à la transition vers la cytokinèse et à la réinitialisation des centrosomes vers un état interphasique.





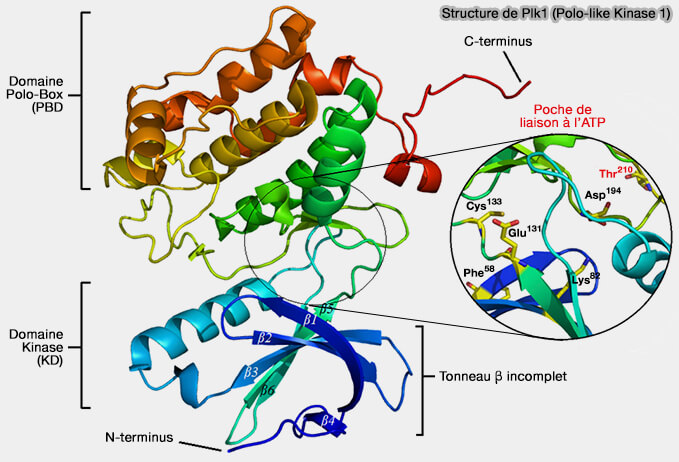
 structure des kinases) et est soumis à une régulation stricte par phosphorylation (Structure of the Catalytic Domain of Human Polo-like Kinase 1 2007).
structure des kinases) et est soumis à une régulation stricte par phosphorylation (Structure of the Catalytic Domain of Human Polo-like Kinase 1 2007).