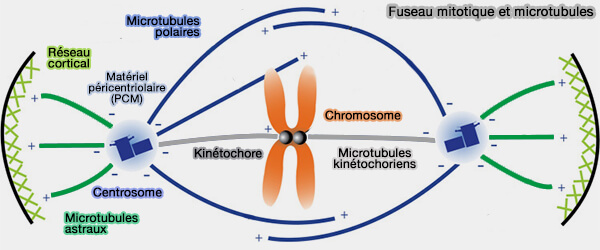
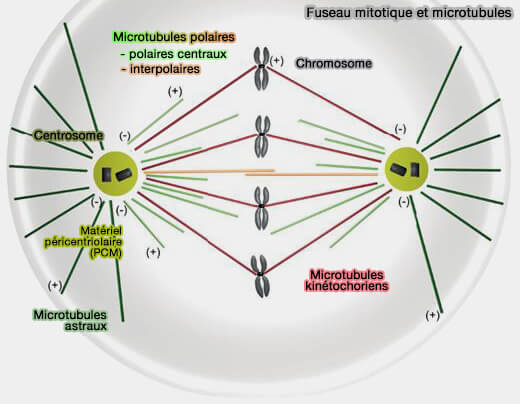
Les microtubules polaires et astraux structurent le fuseau mitotique en générant des forces d’organisation, de positionnement et d’écartement des pôles, sans interaction directe avec les chromosomes.
Fuseau mitotique et microtubules
(Figure : vetopsy.fr d'après Ryniawec et coll)
L’architecture du fuseau repose sur la coexistence de trois grandes populations de microtubules, spatialement et fonctionnellement spécialisées :
Les microtubules polaires émanent des deux pôles du fuseau mitotique et ne s’attachent pas aux kinétochores.
Les microtubules polaires constituent une population hétérogène du fuseau mitotique, comprenant :
d’une part, des microtubules polaires centraux dynamiques, qui contribuent à la densité et à l’équilibre mécanique autour de la plaque équatoriale,
d’autre part, des microtubules interpolaires, organisés en faisceaux antiparallèles dans la zone médiane, responsables du glissement et de l’allongement du fuseau en anaphase B.
Microtubules polaires centraux dynamiques
Les microtubules polaires orientés vers la région centrale (SMT) ne participent pas au chevauchement antiparallèle et ne génèrent pas directement des forces directionnelles,
1. Cette population courte et très dynamique assure un rôle de support mécanique et d’intégration dynamique autour de la plaque équatoriale.
2. Ils contribuent au fonctionnement du fuseau de plusieurs manières complémentaires.
a. Ils augmentent la densité microtubulaire périchromosomique :
en s’étendant vers la région équatoriale sans s’y verrouiller,
en renforçant le maillage microtubulaire autour des chromosomes, ce qui améliore la robustesse mécanique locale du fuseau en métaphase.
b. Ils participent à la répartition spatiale des contraintes mécaniques entre pôles et chromosomes, en amortissant les fluctuations de tension générées par les microtubules kinétochoriens et par les forces d’expansion interpolaire.
c. lls interagissent préférentiellement avec des MAP actives dans la région chromosomique, notamment :
TPX2, qui augmente leur persistance et favorise une orientation compatible avec l’axe du fuseau,
HURP, qui stabilise localement ces microtubules à proximité immédiate des chromosomes,
XMAP215/ch-TOG, qui soutient leur croissance rapide et leur renouvellement.
Ces interactions ne les transforment pas en fibres structurées, mais maintiennent un réseau microtubulaire dense, plastique et réactif, indispensable à l’alignement métaphasique.
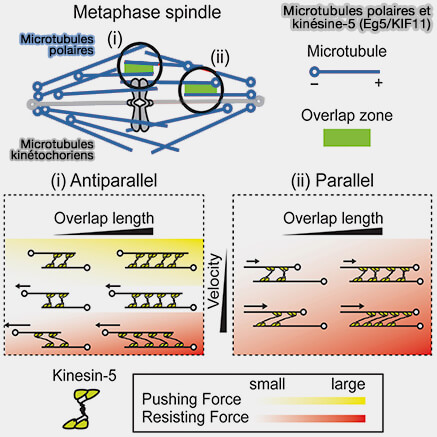
Microtubules interpolaires et zone médiane du fuseau
1. Les microtubules interpolaires ne sont pas attachés aux kinétochores, mais s’organisent dans la région médiane du fuseau en faisceaux antiparallèles, orientés tête-bêche avec leurs extrémités (+) dirigées l’une vers l’autre, via des protéines associées aux microtubules
comme PRC1 et des kinésines de la zone médiane ( kinésines du fuseau).
Microtubules polaires et kinésine-5 (Eg5/KIF11)
(Figure : vetopsy.fr d'après Shimamoto et coll)
Cette organisation antiparallèle définit une zone fonctionnelle distincte de la plaque équatoriale, dépourvue de chromosomes, appelée zone médiane, qui constitue un module mécanique dédié à l’allongement du fuseau et préfigure la formation du corps intermédiaire (midbody) en télophase.
L’architecture du chevauchement est structurée par des protéines associées aux microtubules telles que PRC1, qui stabilisent l’alignement antiparallèle et servent de plateforme de recrutement pour les moteurs et régulateurs de la zone médiane.
2. Les microtubules interpolaires ont pour fonction principale de générer et de transmettre les forces responsables de l’allongement du fuseau et de l’écartement des pôles, contribuant à l’amplification spatiale de la ségrégation chromosomique, sans exercer de traction directe sur les chromosomes.
4. Selon la phase de la mitose, cette architecture peut être mécaniquement contrainte ou extensible, ce qui conditionne la capacité des microtubules polaires à générer un allongement effectif du fuseau.
Les forces internes générées dans la zone médiane sont complétées par la traction exercée sur les microtubules astraux par la dynéine corticale ( force extérieure).
Cette traction, appliquée aux pôles du fuseau, est coordonnée avec l’état mécanique de la zone médiane et contribue, selon le contexte mitotique, soit à la stabilisation de la distance inter-polaire, soit à son augmentation effective.
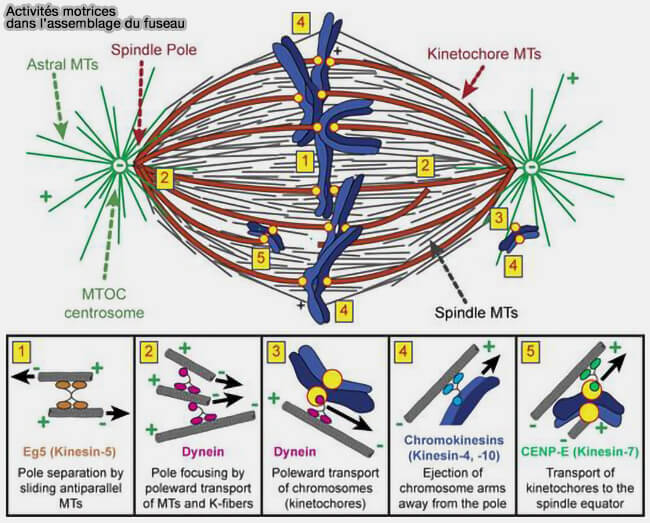
Activités motrices dans l'assemnblage du fuseau
(Figure : vetopsy.fr d'après Heald et Khodjakov)
L’aster désigne l’ensemble de ces microtubules astraux, disposés radialement autour du centrosome, formant une structure étoilée visible surtout en prophase et en prométaphase.
Remarque : la dynamique des microtubules astraux et le positionnement du fuseau sont également modulés par des kinésines associées aux extrémités (+), telles que KIF18B, qui contribuent au positionnement et au centrage du fuseau mitotique (+TIPs spécialisés).
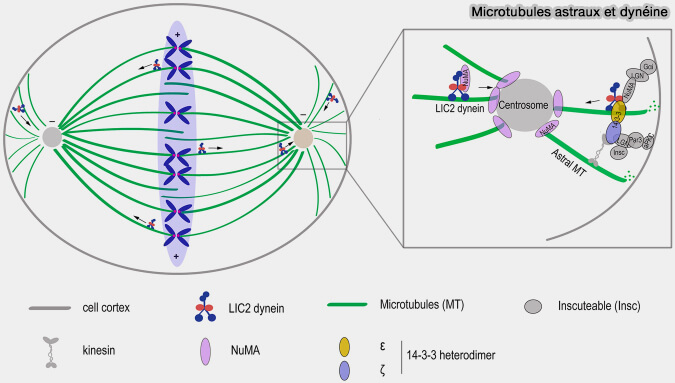
Microtubules astraux et dynéine
(Figure : vetopsy.fr d'après Mahale et coll)
le positionnement spatial correct du fuseau mitotique au centre de la cellule,
l’orientation de l’axe de division en fonction de la géométrie cellulaire et des signaux corticaux,
la stabilisation globale de l’architecture bipolaire du fuseau par des forces de traction externes,
la coordination entre l’organisation du fuseau et la polarité cellulaire en vue de la cytokinèse.
3. Selon la phase de la mitose, la force imposée par la dynéine corticale peut conditionner ou non la capacité des microtubules polaires à générer un allongement effectif du fuseau.
La dynéine corticale est recrutée au cortex cellulaire par le complexe Gαi–LGN–NuMA, qui couple l’activité motrice de la dynéine aux microtubules astraux et permet la génération de forces de traction participant au positionnement et à l’écartement des pôles du fuseau.
a. En prométaphase et en métaphase, bien que les microtubules polaires s’organisent déjà antiparallèlement, la zone médiane reste courte, dense et mécaniquement contrainte. PRC1 stabilise localement le chevauchement sans permettre un glissement extensif, tandis que KIF4A limite la polymérisation des microtubules chevauchants.
Dans ces phases, les forces générées par les moteurs interpolaires et la dynéine sont absorbées par la rigidité centrale du fuseau, verrouillant la distance entre les pôles.
Ce verrouillage mécanique empêche un allongement prématuré du fuseau et permet l’établissement progressif de la biorientation chromosomique avant la transition vers l’anaphase B.
b. En anaphase B, la traction exercée par la dynéine corticale sur les microtubules astraux ne se limite plus à stabiliser la géométrie du fuseau, mais contribue activement à l’écartement des pôles.
En tirant les centrosomes vers la périphérie cellulaire, elle contribue activement à l’allongement du fuseau, en synergie avec le glissement des microtubules polaires antiparallèles




 kinésines du fuseau).
kinésines du fuseau).