La cytokinèse est régulée par des modules équatoriaux spécialisés et par l’activité coordonnée d’Aurora B, des phosphatases PP1/PP2A et des complexes du fuseau central, qui conditionnent l’activation locale de RhoA, la formation du sillon et l’engagement de l’abscission.
Régulations moléculaires de la cytokinèse
1. Les régulations de la cytokinèse sont ici abordées sous un angle fonctionnel et spatial, en mettant l’accent sur les acteurs équatoriaux assurant le couplage entre le fuseau central et le cortex cellulaire.
Les actions d’Aurora B et du complexe CPC, ainsi que celles des phosphatases PP1/PP2A, qui assurent la transition ordonnée entre la mitose nucléaire et la division du cytoplasme, sont abordées dans cette page.
Les rôles de CDK1 et de Plk1 dans la sortie de mitose sont développés dans leurs chapitres correspondants ( sortie de mitose).
2. La mise en place et la dynamique de la cytokinèse reposent sur un réseau dense de régulateurs, illustré notamment par l’identification de plusieurs dizaines de protéines impliquées dans l’assemblage de l’anneau contractile chez la levure, modèle ayant largement contribué à la compréhension des principes généraux de la cytokinèse (Assembly Mechanism of the Contractile Ring for Cytokinesis by Fission Yeast 2008).
2. Ce complexe s’accumule spécifiquement sur les microtubules antiparallèles de la zone médiane du fuseau, puis persiste au niveau du pont intercellulaire et du corps intermédiaire.
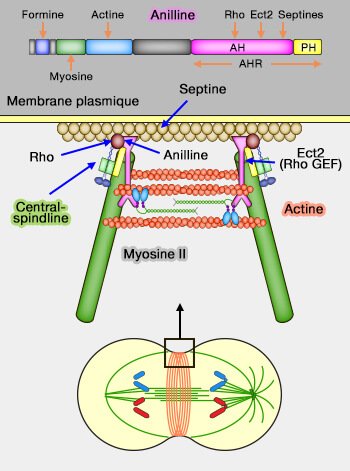
Couplage de l’anneau contractile à la membrane plasmique
Pour que la constriction de l’anneau contractile aboutisse à la séparation des deux cellules filles, la force générée par le réseau actomyosine doit être transmise à la membrane plasmique.
Ce couplage mécanique repose sur plusieurs protéines spécialisées.
a. Sa région C-terminale contient un domaine PH (Pleckstrin Homology), responsable de l’ancrage à la membrane plasmique via les phosphoinositides, condition essentielle à la transmission de la force de constriction.
Les septines forment en effet des structures filamentaires corticales au niveau du sillon de clivage, où elles agissent comme échafaudages mécaniques et barrières de diffusion, stabilisant l’ancrage de l’anneau contractile à la membrane plasmique et confinant la machinerie de cytokinèse à la région équatoriale (Anillin 2010).
En cytokinèse tardive, la région AH de l’anilline peut également contribuer à son association avec les microtubules du pont intercellulaire, renforçant le couplage entre cortex équatorial et architecture microtubulaire, sans jouer un rôle organisateur du fuseau (Anillin directly crosslinks microtubules with actin filaments 2025).
c. La région N-terminale comporte :
une région fonctionnelle associée à la formine,
des domaines d’interaction avec l’actine filamentaire et la myosine II, permettant de stabiliser mécaniquement l’anneau contractile au cortex équatorial.
Ce domaine C1, distinct des domaines C1 classiques des protéines kinases C, ne fonctionne pas comme un capteur de diacylglycérol et n’exerce aucune activité enzymatique, mais contribue à l’ancrage transitoire du complexe centralspindlin à la membrane plasmique, renforçant le couplage entre le fuseau central, le cortex équatorial et l’anneau contractile lors de la cytokinèse.
Abscission
L’abscission correspond à l’étape finale de la cytokinèse et assure la scission membranaire irréversible au niveau du corps intermédiaire (midbody), après l’achèvement de la constriction cytoplasmique.
Elle repose sur la transformation progressive de la zone médiane en une plateforme spécialisée nécssaire à la séparation finale des deux cellules filles.
L’abscission est un événement distinct de la constriction de l’anneau contractile, soumis à un contrôle strict et retardé tant que l’intégrité du matériel génétique n’est pas assurée.
L’abscission marque ainsi la transition finale entre une cellule en division et deux cellules filles autonomes, tant sur le plan structural que fonctionnel.
Contrôle de l’engagement de l’abscission par Aurora B (NoCut)
L’anneau contractile assure l’étranglement du cytoplasme, mais l’abscission ne s’engage pas automatiquement à l’issue du recrutement de la machinerie ESCRT et demeure soumise à un contrôle terminal.
La présence de chromatides retardataires, de ponts chromosomiques (chromatin bridges) ou de défauts de ségrégation persistants bloque la progression vers l’abscission et empêche la coupure membranaire finale.
Tant qu’Aurora B reste active au niveau du corps central, l’abscission est inhibée.
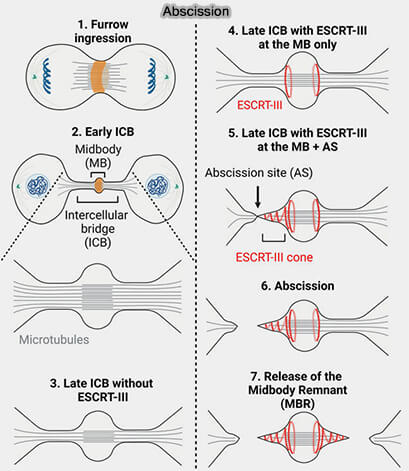
Mécanisme de l'abscission
(Figure : vetopsy.fr d'après Andrade et Echard)
2. Aurora B met en œuvre un mécanisme de contrôle terminal, appelé checkpoint d’abscission (NoCut), qui retarde la coupure membranaire en présence d’anomalies chromosomiques.
3. La levée du checkpoint d’abscission intervient lorsque les anomalies chromosomiques ont été résolues.
Extinction enzymatique du signal Aurora B en cytokinèse tardive
L'extinction du signal Aurora B constitue une étape clé de la cytokinèse tardive, distincte du checkpoint d’abscission proprement dit.
Elle correspond à la neutralisation progressive de l’activité kinase maintenue au niveau du corps intermédiaire et conditionne la levée définitive de l’état pré-abscission.
Cette déphosphorylation provoque la disparition de son gradient équatorial, marquant l’extinction spatiale du signal inhibiteur.
b. Les complexes PP2A-B56 et PP2A-B55 déphosphorylent plusieurs cibles majeures d’Aurora B encore actives en cytokinèse tardive, notamment Nedc80, KNL1, MKLP1/KIF23 et PRC1.
Cette déphosphorylation réduit l’impact fonctionnel du signal Aurora B sur les structures équatoriales et met fin aux contraintes moléculaires maintenant l’état pré-abscission.
2. La disparition du gradient d’Aurora B entraîne une série de transitions moléculaires clés, incluant :
la perte de la stabilisation équatoriale imposée sur centralspindlin et PRC1,
la levée des freins équatoriaux qui maintenaient l’état pré-abscission,
la création d’un état permissif au recrutement séquentiel de la machinerie ESCRT.
Cette extinction enzymatique constitue ainsi le verrou moléculaire reliant la résolution des événements mitotiques à l’engagement irréversible de la coupure membranaire finale.
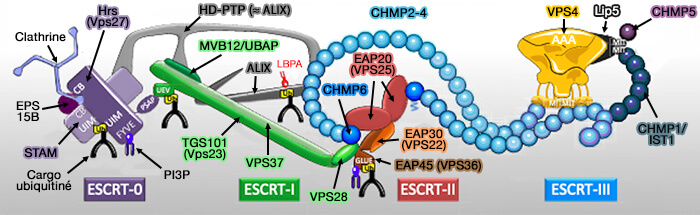
Mécanisme moléculaire de l’abscission
L’exécution de l’abscission correspond à l’étape terminale de la cytokinèse et repose sur l’activation mécanique de la machinerie complexe ESCRT-III (Endosomal Sorting Complex Required for Transport), désormais libérée de l’inhibition imposée par le checkpoint d’abscission et par l’activité résiduelle d’Aurora B.
a. Elle permet le recrutement des adaptateurs ALIX et TSG101 qui assurent le recrutement fonctionnel de la voie ESCRT.
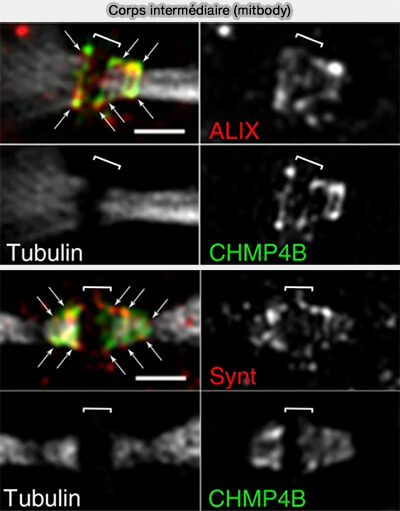
Corps intermédiare (midbody)
(Figure : vetopsy.fr d'après Addi, et coll)
Ces adaptateurs permettent l’organisation locale du complexe ESCRT-III, dont les sous-unités CHMP s’assemblent progressivement au niveau d’un site d’abscission distal, spatialement distinct du centre du midbody.
CHMP4B constitue l’effecteur clé de la constriction membranaire lors de l’abscission.
b. L’assemblage d’ESCRT-III s’accompagne de la polymérisation de filaments hélicoïdaux capables de générer une contrainte de courbure membranaire.
Cette organisation entraîne l’amincissement progressif du pont intercellulaire et prépare la membrane à la fission finale ( mécanisme de ESCRT-III)
2. En parallèle, la spastine, ATPase de type AAA, est recrutée pour assurer la section des microtubules résiduels du corps intermédiaire, levant les dernières contraintes mécaniques opposées à la coupure membranaire.
Remarque : cette cascade fonctionnelle, i.e. PRC1/centralspindlin ➞ CEP55 ➞ ALIX et TSG101 ➞ ESCRT-III (CHMP4B) ➞ spastine ➞ abscission, décrit la continuité moléculaire reliant l’organisation du fuseau central à la coupure membranaire finale.
L’abscission s’achève par la séparation complète et irréversible des deux cellules filles, marquant la fin du processus de division cellulaire.
Contribution des endosomes à l’abscission
1. Les endosomes ou corps multivésiculaires (MVE/MVB) participent activement à l’abscission en fournissant un support membranaire et organisationnel à la machinerie ESCRT, un mode d’action caractéristique de cette voie.
Ils apportent localement des membranes et des lipides au niveau du pont intercellulaire par des événements de fusion contrôlée avec la membrane plasmique, fournissant la surface nécessaire au remodelage et à l’amincissement progressif de la zone d’abscission.
2. En parallèle, les MVB contribuent au recrutement et à l’organisation locale de la machinerie ESCRT en véhiculant des adaptateurs, des protéines de tri et des lipides favorisant la courbure membranaire.
1. La réorganisation du cortex et de la membrane plasmique prépare la stabilisation des deux cellules filles.
Après l’abscission, la membrane plasmique se referme et se stabilise autour de chaque cellule fille.
Le cortex d’actine se réorganise en un réseau isotrope, les tensions équatoriales disparaissent et la mécanique corticale retrouve les propriétés requises pour l’adhésion, la migration et la reprise des fonctions interphasiques.
2. La réactivation progressive des programmes nucléaires marque la sortie définitive de la mitose.
Le réassemblage des pores nucléaires, la réimportation des facteurs nucléaires, la reprise de la transcription et la reconstitution de l’architecture chromatinienne ramènent progressivement les deux noyaux en état fonctionnel de phase G1.
Ce rétablissement coordonné marque la fin de la division cellulaire et la restitution d’un cycle cellulaire opérationnel.




 sortie de mitose).
sortie de mitose).