Aurora B est la kinase centrale du complexe CPC qui assure la correction des attachements kinétochore-microtubules, le contrôle du SAC et l’orchestration tardive de la cytokinèse.
a. Son activité ne déclenche pas l’entrée en mitose, mais s’inscrit en aval du signal complexe cycline B/CDK1 (MPF), qu’elle exploite pour surveiller, corriger et stabiliser les attachements kinétochore-microtubules, ainsi que pour contrôler le point de contrôle du fuseau (SAC), sans intervenir dans l’organisation globale du fuseau mitotique.
de Plk1, qui coordonne de manière globale les transitions mitotiques et l’enchaînement ordonné des événements mécaniques et enzymatiques.
2. Aurora B opère selon une logique chromosomique et tension-dépendante, fondée sur sa localisation dynamique au centromère, au kinétochore interne, puis à la région équatoriale, où son activité est continuellement ajustée par la géométrie du fuseau et l’état des forces mécaniques.
Structure d'Aurora B et du CPC (Chromosomal Passenger Complex)
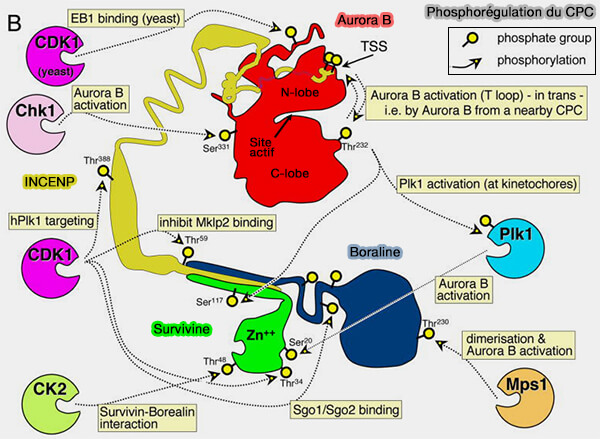
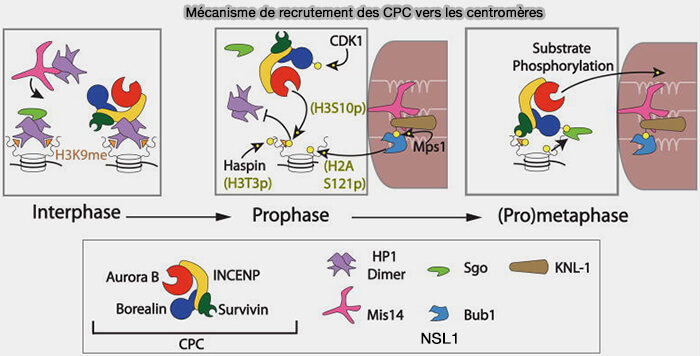
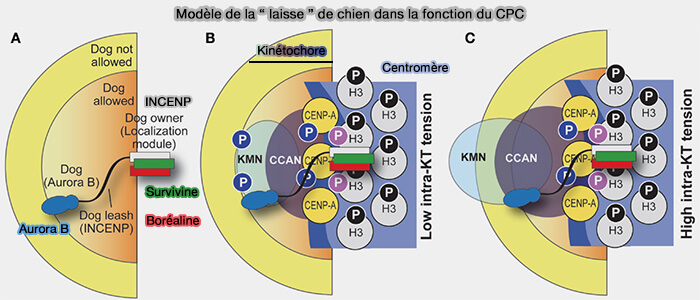
Aurora B n’exerce pas son activité sous forme isolée, mais fonctionne comme la sous-unité catalytique du Chromosomal Passenger Complex (CPC), dont l’organisation structurale, la composition et la dynamique conditionnent directement son activation et ses fonctions mitotiques.
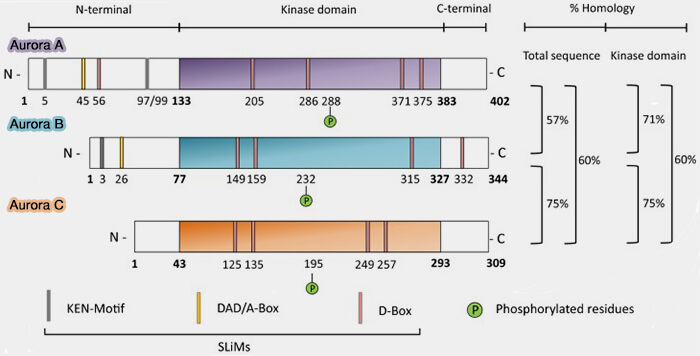
Structure des kinases Aurora
(Figure : vetopsy.fr d'après Willems et coll)
un lobule C-terminal, majoritairement α-hélicoïdal, responsable de l’orientation du substrat et de l’activité catalytique,
une boucle d’activation interlobulaire contenant un résidu thréonine conservé, Thr232 chez l’homme, dont la phosphorylation est indispensable à l’acquisition d’une activité enzymatique efficace.
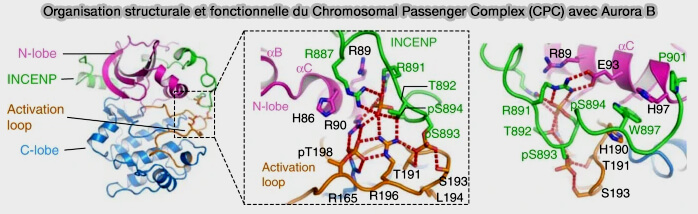
Organisation structurale et fonctionnelle du Chromosomal Passenger Complex (CPC) avec Aurora B
(Figure : vetopsy.fr d'après Azeez et coll)
2. À l’état basal, la boucle d’activation d’Aurora B adopte une conformation instable incompatible avec une catalyse efficace, ce qui limite fortement son activité en l’absence de cofacteur, en particulier INCENP.
3. Le tableau suivant, qui n’a pas vocation à être exhaustif, montre les principaux substrats phosphorylés par Aurora B au cours du cycle mitotique, classés selon la phase mitotique dominante, la localisation subcellulaire et la conséquence fonctionnelle principale, afin de mettre en évidence son rôle transversal.
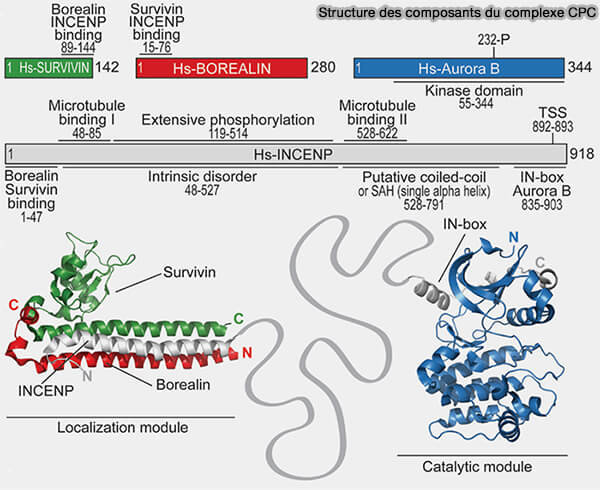
Structure des composants du CPC (Chromosomal Passenger Complex)
(Figure : vetopsy.fr d'après Krenn et Musachio)
2. Les protéines dites " passagères " se caractérisent par leur intégration transitoire au sein du CPC constitué de :
Aurora B, sérine/thréonine kinase, dont l’activité mitotique dépend strictement de son intégration au sein du complexe,
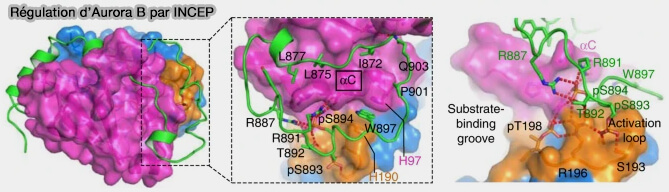
INCENP (INner CENtromere Protein), plateforme d’assemblage et cofacteur allostérique essentiel, assurant l’activation locale d’Aurora B par la stabilisation de sa boucle d’activation, condition nécessaire à une catalyse efficace,
a. La liaison d’INCENP au domaine catalytique d’Aurora B stabilise la boucle d’activation de la kinase dans une conformation compatible avec la catalyse, conditionnant ainsi l’établissement d’un état enzymatique actif.
Cette interaction favorise l’autophosphorylation d’Aurora B et permet un accès efficace aux substrats, sans modifier la spécificité intrinsèque de la kinase.
b. INCENP agit donc comme un coactivateur enzymatique permissif, garantissant que l’activité d’Aurora B ne s’exprime qu’au sein des plateformes structurales où le CPC est correctement assemblé et positionné.
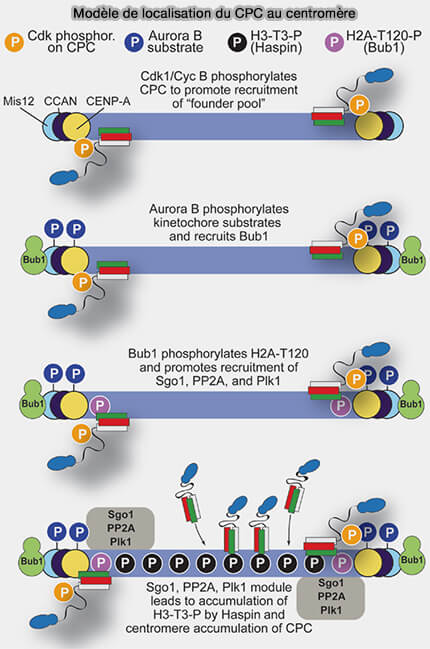
Modèle de localisation du CPC au centromère
(Figure : vetopsy.fr d'après Krenn et Musachio)
1. À l’entrée en mitose, la phosphorylation de l’histone H3 sur Thr3 (H3-Thr3-P) constitue un signal chromatinien clé.
Elle est catalysée par la kinase Haspin activée par des phosphorylations dépendantes du complexe cycline B/CDK1 (MPF) et de Plk1, établissant un lien direct entre l’activation mitotique globale et la signalisation chromatinienne locale.
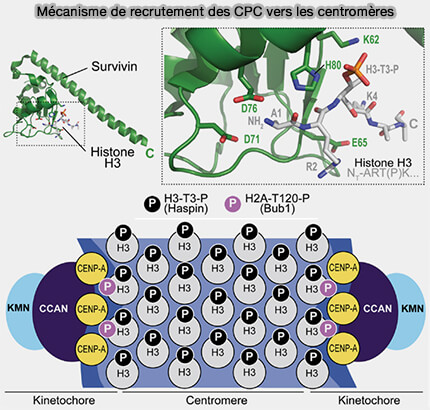
Elle crée un site d’ancrage spécifique reconnu par le complexe CPC via la survivine, permettant le recrutement initial d’Aurora B au centromère interne, préalable à l’établissement du système de correction des attachements kinétochoriens.
Remarque : le terme " centromère interne " est utilisé dans le contexte d’Aurora B et du Chromosomal Passenger Complex (CPC) pour désigner une zone fonctionnelle plutôt qu’une entité structurale distincte.
Il correspond à la chromatine péricentromérique située entre les deux kinétochores sœurs, enrichie en nucléosomes H3/H4 canoniques porteurs de marques mitotiques spécifiques, notamment H3-Thr3-P et H2A-Thr120-P.
Cette région constitue une plateforme chromatinienne de signalisation tension-dépendante, permettant le recrutement et l’activité du CPC au voisinage immédiat des kinétochores, sans correspondre au centromère CENP-A, ni au CCAN (Constitutive Centromere-Associated Network) proprement dits.
2. Ce mécanisme est renforcé par une voie parallèle impliquant la marque chromatinienne H2A-T120-P, histone H2A phosphorylée sur Thr120 par Bub1, qui recrute les shugoshines et contribue à la stabilisation du positionnement du CPC au centromère interne, en renforçant l’ancrage centromérique face aux forces exercées par le fuseau ( rôles des shugoshines).
En reconnaissant H2A-T120-P, les shugoshines établissent une plateforme centromérique stable qui, via le recrutement local de PP2A, module l’état de phosphorylation des composants centromériques.
Recrutement de CPC au centromère par la survivine
(Figure : vetopsy.fr d'après Krenn et Musachio)
Elles contrebalancent les forces de dissociation susceptibles de déplacer le CPC vers le kinétochore externe, assurant ainsi le maintien d’un gradient d’activité d’Aurora B centré sur la chromatine péricentromérique, lien mécanique et signalétique avec le fuseau mitotique ( complexe KMN).
3. L’intégration de ces signaux assure un ciblage précis du CPC au centromère interne, condition indispensable à l’activité correctrice d’Aurora B sur les attachements kinétochoriens.
Elle permet en parallèle la stabilisation sélective des attachements soumis à une tension bipolaire adéquate, préalable à la satisfaction du point de contrôle du fuseau (SAC).
Cette transition ne déclenche pas à elle seule la migration du CPC, mais lève les contraintes qui maintenaient Aurora B et ses partenaires au centromère.
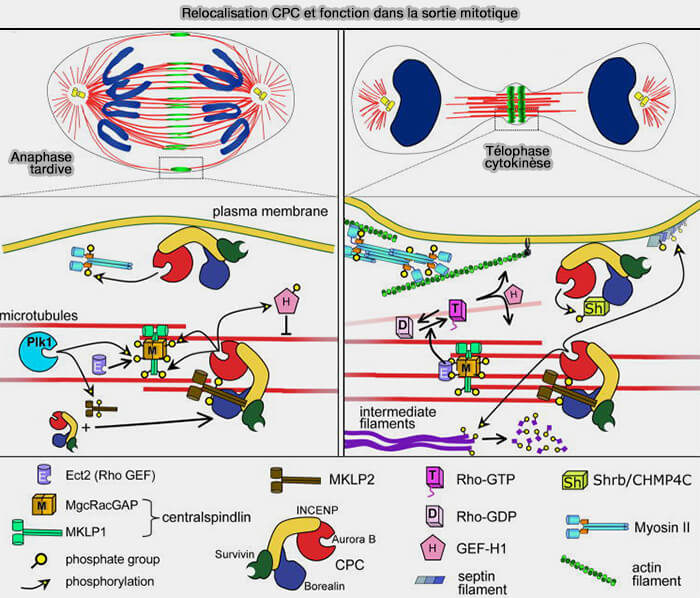
Cette redistribution devient pleinement visible et fonctionnellement stabilisée en télophase, lorsque le CPC se concentre dans la zone équatoriale (plaque équatoriale), puis sur le fuseau central et enfin au niveau du futur sillon de division et du corps central.
b. La déphosphorylation d'INCENP par Cdc14, phosphatase activée en sortie de mitose, préalablement phosphorylée par CDK1, consolide l’ancrage du complexe sur le fuseau central.
Remarque : la phosphatase Cdc14 joue un rôle central et indispensable dans la sortie de mitose chez la levure, où elle gouverne la bascule globale kinases-phosphatases.
Chez les mammifères, ses homologues CDC14A et CDC14B sont conservés mais n’assurent pas cette fonction maîtresse.
Leur action est restreinte, locale et contextuelle, notamment au niveau du CPC et d’INCENP lors du repositionnement tardif du complexe sur le fuseau central.
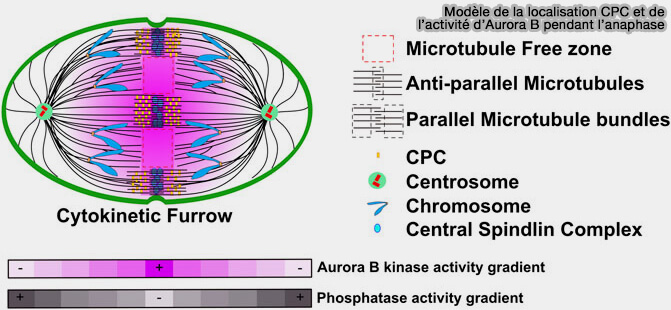
Établissement du gradient équatorial d’Aurora B
Une fois le CPC stabilisé sur le fuseau central, Aurora B établit un domaine équatorial d’activité kinase.
Aurora B et phosphatases pendant l'anaphase
(Figure : vetopsy.fr d'après Ali et Stukenberg)
1. Son activité est maximale au centre du fuseau et décroît progressivement vers le cortex équatorial, générant un gradient de phosphorylation orienté perpendiculairement à l’axe du fuseau, maximal au niveau du fuseau central et décroissant vers la périphérie cellulaire.
2. Ce gradient, structuré par la stabilisation du CPC via MKLP2 et ses interactions avec le réseau d’actomyosine, s’étend jusqu’au cortex équatorial, où l’activité kinase d’Aurora B devient faible mais fonctionnellement significative.
Dans cette région corticale, l’activité résiduelle d’Aurora B est suffisante pour moduler l’état de phosphorylation des myosines corticales et des régulateurs associés du cortex actomyosine.
Cette modulation définit une zone corticale contractile correspondant au futur sillon de clivage, spatialement alignée avec le plan équatorial du fuseau.
3. Le signal équatorial ainsi généré fournit l’instruction positionnelle reliant la géométrie du fuseau central à l’activation localisée des effecteurs de la cytokinèse, notamment centralspindlin et les régulateurs de la voie RhoA.
Transmission du signal équatorial
d’Aurora B au complexe centralspindlin
L’établissement du domaine équatorial d’activité d’Aurora B sur le fuseau central permet le recrutement sélectif du complexe centralspindlin dans la zone médiane.
MKLP1 et MgcRacGAP forment le complexe hétérotétramérique constitutif qui est préexistant.
L'extinction du complexe cycline B/CDK1 (MPF) lève l’inhibition mitotique sur Plk1, qui devient dominant, permettant la phosphorylation de MKLP2, ce qui autorise la relocalisation et le maintien du CPC au niveau équatorial.
CDK1 inhibe fonctionnellement PLK1 en maintenant les substrats et les plateformes de la cytokinèse dans un état phosphorylé et spatialement inactif, empêchant PLK1 d’exercer ses fonctions tardives tant que la métaphase n’est pas achevée.
Remarque : ce positionnement s’appuie sur l’architecture des microtubules antiparallèles, organisée et stabilisée par PRC1, qui constitue une plateforme structurale essentielle du fuseau central.
1. À partir de l’anaphase, une fois positionné dans la zone équatoriale, le CPC et donc Aurora B, phosphoryle MKLP1 (KIF23) au sein du complexe centralspindlin.
a. Cette phosphorylation :
augmente l’affinité de MKLP1 pour les microtubules antiparallèles,
stabilise centralspindlin sur le fuseau central,
favorise son enrichissement spécifique dans la zone médiane.
b. À ce stade, centralspindlin constitue une plateforme structurale reliant le fuseau central au cortex équatorial, jouant un rôle principalement organisationnel et préparatoire, en amont de l’activation des voies contractiles responsables de la formation et de la progression du sillon.
Centralspindlin est alors stabilisé par son organisation hétérotétramérique associant MKLP1 et MgcRacGAP.
Relocalisation du CPC (Chromosomal Passenger Complex)
(Figure : vetopsy.fr d'après Carmena et coll)
Cette étape ne déclenche pas directement la formation du sillon de clivage.
Elle agit comme un préconditionnement cortical, en rendant le cortex équatorial globalement permissif à l’activation de RhoA, sans imposer encore de focalisation spatiale stricte.
Contrairement à RhoGEF-H1, ECT2 agit comme une GEF effectrice focalisée car son association à centralspindlin restreint spatialement son activité au cortex équatorial situé en regard du fuseau central.
Cette organisation assure une activation locale, soutenue et polarisée de RhoA, condition indispensable à l’assemblage de l’anneau contractile et à la formation puis à la progression du sillon de clivage.
Rôles d'Aurora B pendant le cycle cellulaire
En interphase, Aurora B est présente sous une forme peu active et ne remplit pas de fonction mitotique et son activation et sa spécialisation fonctionnelle sont strictement restreintes aux phases mitotiques.
Aurora B et prophase
En prophase, Aurora B, au sein du complexe CPC (Chromosomal Passenger Complex), s’accumule progressivement sur les chromosomes et les régions centromériques, où elle participe à la mise en place de l’état mitotique, sans intervenir encore dans l’organisation du fuseau.
1. À ce stade, son activité cible principalement des substrats chromatiniens et centromériques, notamment les histones et des composants structuraux du centromère, contribuant à :
la structuration fonctionnelle précoce du centromère,
la mise en place du cadre spatial dans lequel s’exercera ultérieurement le contrôle des attachements kinétochoriens.
2. Cette localisation et cette activité préparent le dispositif correcteur qui sera pleinement mobilisé en prométaphase pour le contrôle des attachements chromosome-microtubules, sans intervention directe encore sur le fuseau mitotique, ni sur la signalisation du point de contrôle du fuseau (SAC).
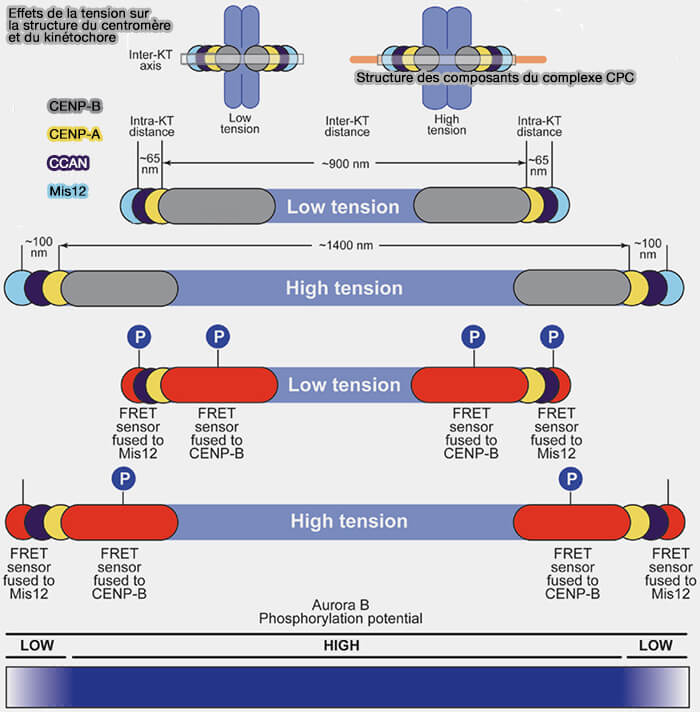
1. Sur le plan mécanistique, Aurora B perçoit l’absence ou l’insuffisance de tension entre les kinétochores sœurs, situation caractéristique des attachements incorrects.
Aurora B phosphoryle des composants du complexe KMN d’attachement kinétochore-microtubule, principalement Ndc80, KNL1 et Dsn1 (complexe Mis12), ce qui réduit l’affinité des microtubules pour les kinétochores lorsque les forces de tension sont insuffisantes ou mal orientées.
Cette activité maintient les attachements incorrects dans un état instable, favorisant leur dissociation et leur correction avant l’anaphase, notamment dans le cas d’attachements syntéliques ou mérotéliques, tant que la tension bipolaire adéquate n’est pas établie.
Tant que la tension exercée entre les chromatides sœurs est insuffisante, Aurora B conserve ainsi une activité corrective.
Effets de la tension sur la structure du centromère et du kinétochore
(Figure : vetopsy.fr d'après Krenn et Musachio)
2. Ce mécanisme de correction assure que seuls les attachements soumis à une tension bipolaire adéquate sont stabilisés, sous la forme d’attachements end-on durables, condition indispensable à l’extinction du point de contrôle du fuseau (SAC) et à la transition vers la métaphase, sans que la correction active ne se poursuive au-delà de cette phase.
Aurora B et métaphase
En métaphase, Aurora B demeure localisée au centromère et au kinétochore interne, au sein du complexe CPC, mais n’agit plus comme une kinase correctrice active. S
Son rôle devient essentiellement permissif et référentiel, dépendant de l’état mécanique atteint par les chromosomes.
1. Lorsque les chromosomes sont correctement bi-orientés et soumis à une tension adéquate, les chromatides soeurs s'éloignent physiquement de la zone centromérique enrichie en Aurora B et en ses substrats kinétochoriens.
Cet éloignement spatial réduit l’accès effectif des composants du complexe KMN à la zone d’activité kinase d’Aurora B, sans nécessiter d’extinction enzymatique active.
2. La diminution de l’influence d’Aurora B permet alors la déphosphorylation relative des composants kinétochoriens par l’action dominante des phosphatases locales, conduisant à la stabilisation passive des attachements microtubule-kinétochore corrects.
Ces attachements end-on stables assurent le maintien de la plaque équatoriale et la persistance d’un état mécaniquement équilibré du fuseau.
3. Aurora B agit ainsi en métaphase comme un capteur mécanosensible spatial, assurant que seuls les attachements soumis à une tension bipolaire adéquate persistent avant la levée du point de contrôle du fuseau (SAC) et la transition vers l’anaphase.
Aurora B en anaphase
En anaphase, Aurora B cesse progressivement son rôle de correcteur des attachements, mais conserve une fonction organisationnelle essentielle au niveau du fuseau central, consécutive à son repositionnement spatial.
2. Son activité contribue à maintenir une organisation contrôlée du midbody, sans intervenir dans les forces motrices d’écartement, mais en assurant la cohérence structurale et temporelle de la division tardive, en particulier jusqu’à l’abscission.
3. Elle participe aussi au contrôle de progression, appelé NoCut checkpoint, qui assure que la séparation chromosomique est entièrement achevée avant l’abscission finale.
Ce mécanisme de surveillance tardif intervient en télophase et au début de la cytokinèse pour détecter la présence éventuelle de chromatides retardataires ou de ponts chromosomiques persistant dans la zone médiane ou au niveau du corps intermédiaire (midbody).
En cas d’ADN résiduel au niveau du corps intermédiaire, l’activité persistante d’Aurora B au midbody empêche l’engagement final de l’abscission, retardant la cytokinèse jusqu’à résolution complète de l’anomalie.
L’extinction progressive de l’activité d'Aurora B ne constitue pas un mécanisme autonome, mais s’inscrit dans la bascule globale kinases-phosphatases qui gouverne la sortie de mitose, conduisant au démantèlement du fuseau tardif, à la transition vers la cytokinèse et à la réinitialisation des centrosomes vers un état interphasique.
Aurora B en cytokinèse et lors de l'abscission
En cytokinèse, Aurora B agit comme un régulateur temporel et spatial de la séparation cellulaire en contrôlant l’engagement et le déroulement de l’abscission.




 structure des kinases),
structure des kinases),