


Cycle cellulaire
Reproduction cellulaire
Méiose (reproduction sexuée)
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

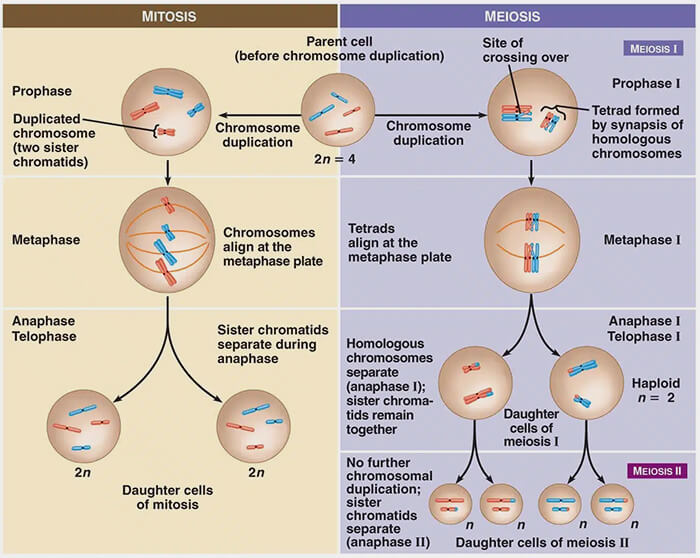
La méiose est une division cellulaire spécialisée qui produit des cellules haploïdes, les gamètes (spermatozoïdes et ovules), par une succession de deux divisions, reposant sur des mécanismes distincts de la mitose.
Principes généraux de la méiose
1. La méiose est un processus de division cellulaire propre aux cellules germinales, dont la fonction est de réduire de moitié le nombre de chromosomes et de générer une diversité génétique au sein des gamètes (spermatozoïdes et ovules).
Les cellules germinales se distinguent des cellules somatiques par leur destinée reproductive.
- Elles constituent une lignée cellulaire spécialisée, séparée très précocement au cours du développement embryonnaire, dont la fonction exclusive est de produire des gamètes.
- À l’inverse, les cellules somatiques forment l’ensemble des cellules différenciées de l’organisme, participent à la constitution des tissus et des organes, et se divisent uniquement par mitose sans modifier le nombre de chromosomes transmis.
2. La méiose conduit à la formation de cellules haploïdes (n chromosomes), à partir d’une cellule diploïde (2n chromosomes), par l’enchaînement de deux divisions successives précédées d’une seule phase de réplication de l’ADN ( phase S).
phase S).
Cette réduction chromosomique est indispensable au maintien du caryotype de l’espèce au cours des cycles de fécondation, la fusion de deux gamètes haploïdes restaurant l’état diploïde du zygote.
3. Au-delà de la réduction du nombre de chromosomes, la méiose introduit une variabilité génétique majeure par deux mécanismes complémentaires :
- le brassage interchromosomique, lié à l’orientation aléatoire des paires de chromosomes homologues,
- le brassage intrachromosomique, résultant des échanges de segments entre chromatides non sœurs de chromosomes homologues lors de la recombinaison.

La méiose n’est pas une simple variante de la mitose, mais une division spécialisée adaptée à la production de gamètes, combinant réduction chromosomique et diversification génétique.
(Figure : vetopsy.fr d'après schoolworkhelper.net)
Bases mitotiques communes à la méiose
1. Sur le plan structural et enzymatique, la méiose réutilise l’essentiel de l’appareillage mitotique classique.
- Le fuseau de division est organisé à partir des centrosomes et repose sur un réseau dynamique de microtubules kinétochoriens, polaires et astraux, dont la mécanique générale est comparable à celle observée en mitose.
- Les chromosomes sont reliés au fuseau par des kinétochores assemblés sur les centromères, et leur mouvement dépend des mêmes principes fondamentaux de capture, de tension et de forces motrices ( différentes phases de la mitose).
- Les grandes familles d’enzymes mitotiques sont également mobilisées, notamment les complexes cycline/CDK, Plk1, les kinases Aurora A ainsi qu'Aurora B au sein du CPC (Chromosomal Passenger Complex) et les phosphatases PP1 et PP2A qui orchestrent les transitions de phase et la sortie de division.
- Un point de contrôle du fuseau (SAC) est présent en méiose, assurant que la ségrégation chromosomique ne s’engage qu’après l’établissement d’interactions correctes entre chromosomes et fuseau.
2. Cependant, si les composants sont en grande partie partagés avec la mitose, leur organisation fonctionnelle, leur régulation et leurs cibles diffèrent profondément, en particulier lors de la première division méiotique.
Organisation générale et chronologie de la méiose
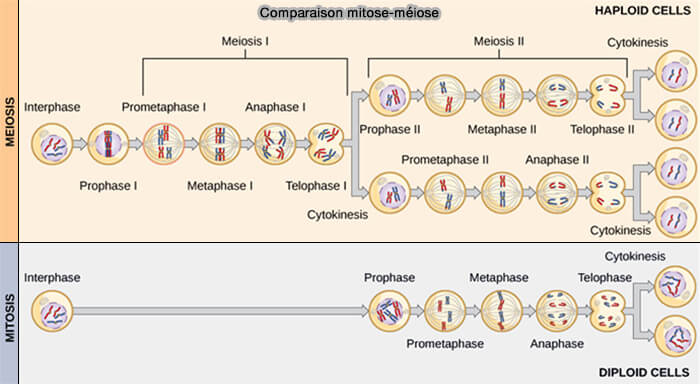
1. Comme en mitose, la méiose, précédée de l'interphase, phase préparatoire du cycle cellulaire au cours de laquelle la cellule croît, réplique son ADN et prépare tous les éléments nécessaires à la division, est divisée en trois phases.
- La phase G1 (de l'anglais growth, croissance ou gap, intervalle ou espace) est la première phase de croissance cellulaire (2n, 2c).
- La phase S (de l'anglais synthesis, synthèse) correspond à la réplication complète du matériel génétique (2n, 4c).
- La phase G2 est la dernière étape de l’interphase, qui a pour fonction principale de préparer la cellule à la méiose en vérifiant la complétude et l’intégrité du génome, tout en assurant la mise en place des structures nécessaires à la division cellulaire (2n, 4c).
(Figure : vetopsy.fr d'après courses.lumenlearning.com)
2. La méiose comprend ensuite deux divisions successives :
- la méiose I dite réductionnelle, qui assure la réduction du nombre de chromosomes,
- la méiose II dite équationnelle sans réplication intermédiaire de l’ADN, qui permet la séparation des chromatides sœurs.
Méiose I dite réductionnelle
1. La méiose I constitue une division fondamentalement non mitotique, bien qu’elle mobilise un appareillage moléculaire en grande partie commun avec la mitose.
Elle est définie par trois propriétés structurantes :
- elle est réductionnelle,
- elle repose sur l’appariement et la recombinaison des homologues,
- elle suspend temporairement la règle mitotique de séparation des chromatides soeurs.
2. La méiose I, qui se déroule en plusieurs phases successives, correspond ainsi à la séparation des chromosomes homologues et constitue l’étape déterminante de la division méiotique.
Prophase I
La prophase I correspond à une phase prolongée au cours de laquelle se mettent en place l’appariement des chromosomes homologues, la synapsis et la recombinaison génétique.
Elle est classiquement subdivisée en plusieurs stades morphologiques, qui correspondent à des étapes fonctionnelles successives.
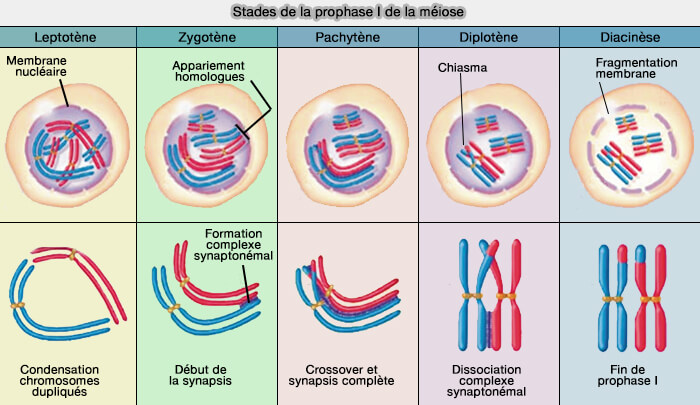
Stade leptotène
Le stade leptotène (du grec, leptos, mince et taenia, ruban) correspond à la condensation initiale des chromosomes répliqués (2n, 4c).
À ce stade, les chromosomes deviennent individualisés dans le noyau et s’engagent dans un processus de reconnaissance de leurs homologues, sans appariement encore établi.
(Figure : vetopsy.fr)
Stade zygotène
Le stade zygotène (du grec, zygos, joug) marque :
- la mise en contact progressive des chromosomes homologues,
- l'initiation de la synapsis.
La synapsis (du grec synápsis, " jonction ", " mise en contact ") désigne l’établissement d’un alignement étroit et continu entre chromosomes homologues tout au long de leur longueur.
1. À partir du stade zygotène, les chromosomes répliqués en phase S (2n, 4c) ne se comportent plus comme des entités indépendantes, comme c’est le cas en mitose.
Alors qu’en mitose chaque chromosome dupliqué interagit de manière autonome avec le fuseau mitotique, la méiose I impose une reconnaissance spécifique entre chromosomes homologues.
a. Cet appariement, absent en mitose, repose sur une reconnaissance homologue précise et conduit à l’alignement des chromosomes sur toute leur longueur.
b. L’appariement constitue une condition indispensable :
- à la recombinaison génétique,
- à la stabilité mécanique des homologues,
- à leur ségrégation correcte lors de l’anaphase I.
2. Cet appariement repose sur la mise en place d’une structure protéique spécialisée, le complexe synaptonémal, qui s’établit progressivement au cours de la prophase I.
La synapsis permet :
- la reconnaissance spécifique et stable des chromosomes homologues,
- leur stabilisation mécanique en tant que paire fonctionnelle,
- la mise en place d’un cadre structural indispensable aux échanges génétiques entre chromatides non sœurs de chromosomes homologues.
L’appariement des homologues constitue ainsi un prérequis absolu à la division réductionnelle, sans équivalent en mitose.
Stade pachytène
Le stade pachytène (du grec, packhus, épais) voit :
- la synapsis complète, appariement strict des chromosomes homologues,
- la recombinaison génétique entre chromatides non sœurs de chromosomes homologues.
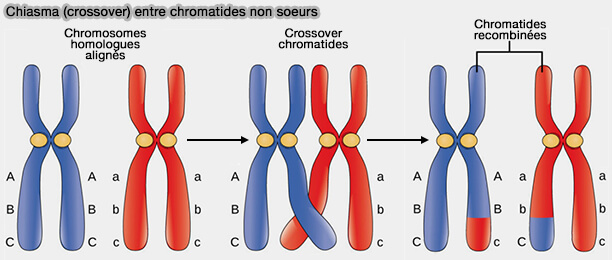
Au sein des paires de chromosomes homologues appariés, des échanges réciproques de segments d’ADN se produisent entre chromatides appartenant à deux chromosomes homologues distincts, et non entre chromatides sœurs d’un même chromosome.
(Figure : vetopsy.fr d'après courses.lumenlearning.com)
1. Ces événements de recombinaison, initiés par des cassures double brin programmées, conduisent à la formation de chiasmas, structures visibles correspondant aux sites d’échange ( crossing-over).
Les chiasmas remplissent une double fonction.
- Ils assurent le brassage génétique intrachromosomique, en combinant des segments issus des deux chromosomes homologues.
- Ils établissent un lien physique entre homologues, indispensable à leur alignement et à leur ségrégation correcte sur le fuseau de méiose I.
2. Ainsi, la recombinaison n’est pas un simple mécanisme de diversification génétique, mais un élément mécanique central de la ségrégation méiotique.
Stade diplotène
Le stade diplotène (du grec, diploos, double) correspond à une séparation partielle des chromosomes homologues, tout en maintenant leur association au niveau des chiasmas formés lors de la recombinaison.
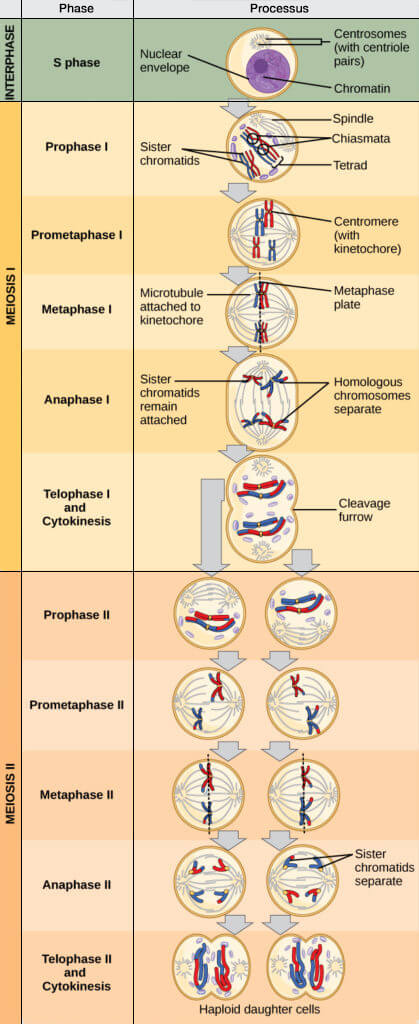
(Figure : vetopsy.fr d'après courses.lumenlearning.com)
Les chiasmas, formés lors de la recombinaison au pachytène, deviennent morphologiquement visibles au diplotène et assurent le maintien d’un lien mécanique entre homologues jusqu’à leur séparation ultérieure.

Chez l’ovocyte, ce stade de diplotène peut durer de nombreuses années, les cellules restant bloquées à ce stade jusqu’au pic de LH déclenchant la reprise de la méiose (maturation du/des ovules dominants).
Stade diacinèse
La diacinèse (du grec, dia, à travers et kinesis, mouvement) correspond :
- à la condensation chromosomique maximale,
- à la préparation de leur engagement sur le fuseau de méiose I.
Elle marque la transition entre les événements nucléaires de la prophase I et l’organisation du fuseau en vue de la métaphase I.
Métaphase I
En métaphase I, les chromosomes homologues s’alignent sur la plaque équatoriale selon une organisation spécifique, distincte de celle observée en mitose, qui prépare directement la ségrégation réductionnelle de l’anaphase I.
Organisation spécifique
de la ségrégation en méiose I
1. En méiose I, la cohésion chromosomique ne remplit pas la même fonction qu’en mitose et obéit à une logique différente.
En mitose, la cohésion entre chromatides sœurs est maintenue jusqu’à la métaphase, puis levée simultanément sur toute la longueur des chromosomes à l’anaphase, permettant leur séparation.
2. Les chromatides sœurs restent solidaires tout au long de la première division tandis que la séparation à venir concerne exclusivement les chromosomes homologues.
a. Cette ségrégation différentielle repose sur :
- la cohésion le long des bras chromosomiques est progressivement levée, ce qui permettra la séparation des chromosomes homologues lors de l’anaphase I,
- la cohésion centromérique entre chromatides sœurs est maintenue.
b. À ce stade, la cellule est toujours en état (2n, 4c.)
3. Parallèlement, l’orientation des kinétochores diffère fondamentalement de celle de la mitose.
En mitose, chaque chromatide sœur possède un kinétochore orienté vers un pôle oppsé du fuseau, ce qui permet leur séparation directe à l’anaphase et conduit à deux cellules filles conservant un état diploïde.
En méiose I, les kinétochores des deux chromatides sœurs d’un même chromosome fonctionnent de manière coordonnée et s’orientent vers un même pôle du fuseau.
Cette co-orientation empêche toute séparation des chromatides sœurs en anaphase I et rend possible la séparation exclusive des chromosomes homologues.
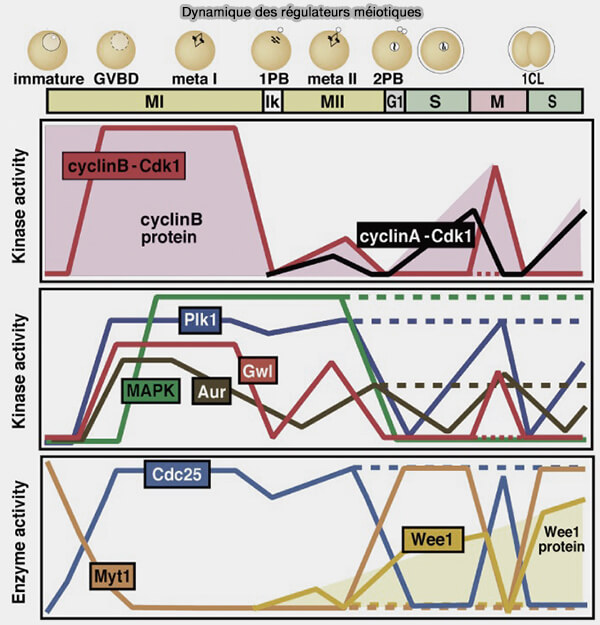
(Figure : vetopsy.fr d'après Kishimoto)
Remarque : dans cette figure, GWL (Greatwall kinase, aussi appelée MASTL chez les mammifères) intervient comme un facteur de stabilisation de l’état mitotique en inhibant la phosphatase PP2A-55B via ENSA/ARPP19, ce qui empêche la déphosphorylation des substrats du complexe cycline B/CDK1 (MPF-based meiotic cell cycle control: Half a century of lessons from starfish oocytes 2018).
Rôle mécanique des chiasmas en métaphase I
1. La stabilité de cette organisation dépend du chiasma, issu d’un crossing-over préalable.
a. Ce lien est stabilisé par la cohésion des chromatides sœurs autour du site d’échange, principalement assurée par les complexes de cohésines méiotiques (notamment REC8), qui maintiennent l’intégrité structurale du couple de chromosomes jusqu’à la métaphase I.
b. Les forces de traction exercées sur les chromosomes homologues proviennent de la dynamique des microtubules (polymérisation/dépolymérisation) et de l’activité des moteurs du fuseau ( forces mécaniques au niveau du fuseau).
- Les kinésines interpolaires Eg5/KIF11 et KIF15 contribuent à l’organisation et à la tension du fuseau bipolaire.
- Au niveau des kinétochores, la kinésine CENP-E/KIF10 participe à la transmission des forces.
- La dynéine kinétochorienne contribue aux mouvements initiaux des chromosomes vers les pôles.
c. La stabilité des attachements est contrôlée par des régulateurs tels que Aurora B (correction tension-dépendante des attachements) et la kinésine dépolymérase MCAK/KIF2C, qui élimine les attachements incorrects.
2. Ce lien mécanique n’est toutefois pas rompu par la force du fuseau, mais est levé par un mécanisme moléculaire strictement contrôlé.
- La séparation des chromosomes homologues en anaphase I résulte du clivage de la cohésine méiotique REC8 sur les bras chromosomiques par la séparase.
- La cohésion centromérique est, quant à elle, protégée par la shugoshine (SGO2) et PP2A, ce qui empêche la séparation des chromatides sœurs à ce stade.
En l’absence de chiasma, cette transmission mécanique ne peut pas s’établir :
- les deux homologues se comportent comme des unités indépendantes,
- les tensions sont mal réparties,
- les attachements kinétochoriens deviennent instables ou aberrants,
- le point de contrôle du fuseau (SAC) est contourné ou insuffisant, ce qui conduit à une augmentation marquée des erreurs de disjonction en anaphase I.
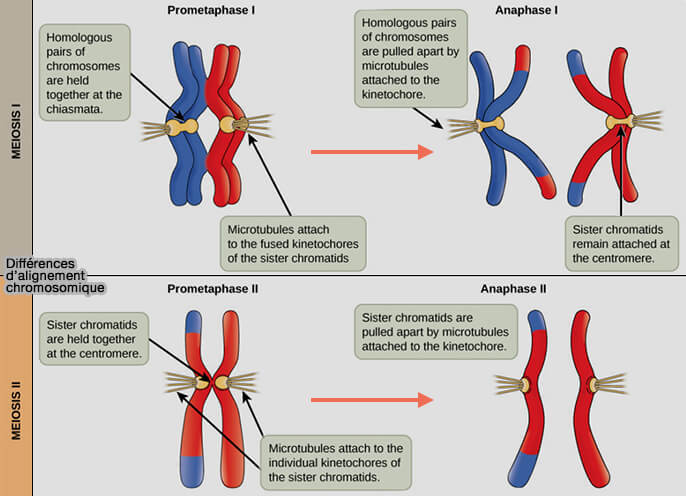
(Figure : vetopsy.fr d'après courses.lumenlearning.com)
Anaphase I
L’anaphase I correspond à la séparation effective des chromosomes homologues, tandis que les chromatides sœurs restent associées jusqu’à la méiose II.
À l’issue de l’anaphase I et de la télophase I, la cellule passe ainsi d’un état (2n, 4c) à un état (n, 2c), chaque chromosome étant encore constitué de deux chromatides sœurs, qui ne seront séparées qu’au cours de la méiose II.
Télophase I et état chromosomique intermédiaire
La télophase I correspond à la fin de la division réductionnelle.
Elle marque l’entrée dans un état chromosomique intermédiaire, qui prépare directement la méiose II.
Méiose II dite équationnelle
La méiose II est une division équationnelle comparable à une mitose classique, mais s’exerce sur un jeu chromosomique haploïde.
En effet, elle n'est pas précédée par une interphase avec duplication des chromosomes en phase S.
Elle comprend les mêmes phases :
- la prophase II, généralement brève, les chromosomes étant restés partiellement condensés,
- la métaphase II, marquée par l’alignement des chromosomes sur la plaque équatoriale,
Lors de l'ovogenèse, c'est l'ovocyte II, arrêté dans son évolution à la métaphase de la méiose II, qui est expulsé lors de l'ovulation.
- l'anaphase II, correspondant à la séparation des chromatides sœurs.
- la télophase II, marquée par la reformation des enveloppes nucléaires autour des lots chromosomiques séparés et la décondensation progressive des chromosomes,
- la cytokinèse et l'abcission, qui assurent la séparation physique des cellules filles et conduisent à la formation de quatre spermatozoïdes (1n, 1c) ou d’un ovule (1n, 1c) accompagné d'un globule polaire (1n, 1c).
Dimorphisme sexuel de la méiose
La méiose présente un dimorphisme sexuel marqué.
La gamétogenèse, ovogenèse et spermatogenèse, est étudiée longuement dans des chapitres spécifiques.
Chez la femelle
1. Le premier arrêt prolongé en diplotène de la prophase I persiste jusqu’à la puberté ( physiologie de la puberté).
a. Celle-ci met en place l’activité cyclique de l’axe hypothalamo-hypophyso-ovarien, rendant possible, à chaque cycle, la reprise transitoire de la méiose lors du pic de LH précédant l’ovulation (maturation du/des ovules dominants).
Durant cette phase prolongée, le complexe synaptonémal a déjà été dissous et la stabilité des paires de chromosomes homologues appariés repose alors uniquement sur la persistance des chiasmas et sur la cohésion des chromatides sœurs.
b. Cet arrêt prolongé favorise la perte progressive de cohésion entre chromatides sœurs, l’altération des complexes de cohésines et une instabilité des chiasmas.
Ces altérations constituent la cause majeure des aneuploïdies, comme la trisomie 21 (syndrome de Down), dont l’incidence augmente fortement avec l’âge maternel.
2. Un second arrêt s’établit ensuite en métaphase II et ne sera levé qu’en cas de fécondation.
Remarque : le stock d’ovocytes étant limité dès la naissance, son épuisement progressif conduit à la ménopause.
Chez le mâle
Chez le mâle, au contraire, la méiose débute à la puberté et se poursuit ensuite de manière continue tout au long de la vie, avec un renouvellement constant des cellules germinales et un taux d’erreurs globalement plus faible.
Retour vers la reproduction cellulaire
Biologie cellulaire et moléculaireConstituants de la celluleReproduction cellulaireMatériel génétiqueFuseauCycle cellulaireInterphasePhase G1Phase SPhase G2MitoseProphasePrométaphaseMétaphaseAnaphaseTélophaseCytokinèseAbcissionMéioseRégulation du cycle cellulaireCyclinesCDKComplexes Cyclines/CDKPoints de contrôles (checkponts)Enzymes mitotiquesMoteurs mitotiquesComplexes SMC (condensines et cohésines)BiochimieTransport membranaire Moteurs moléculairesVoies de signalisation