Physiologiquement, la sensation du goût est provoquée
par la stimulation des récepteurs sensoriels, les bourgeons du goût, localisés dans la cavité
buccale.
Ces cellules gustatives doivent
transformer le signal chimique de la saveur en un signal électrique ;
1. la dissolution de la substance chimique, dans ce cas on parle de substance sapide, dans la salive, 2. sa diffusion vers le pore
gustatif,
3. l'absorption par les microvillosités des cellules
gustativesqui, comme toutes les cellules, sont polarisées.
Les stimuli provoquent une dépolarisation de la cellule par deux grands types de récepteurs :
La dépolarisation provoque la libération d'un neurotransmetteur qui transmet l'excitation aux terminaisons nerveuses, où elle donne
naissance à des potentiels
d'action.
Cette
étape de codage donne une image sensorielle particulière.
1. Le codage est quantitatif : plus la
concentration en molécules sapides augmente (plus les potentiels
d'action sont fréquents), plus la saveur est intense.
2. Le codage qualitatif est moins précis : les fibres gustatives sont sensibles
à toutes les saveurs.
On
localise souvent la perception d'une saveur dans une région précise
de la langue. Ceci est faux car chaque saveur est perçue sur toute
la surface de la langue, mais de manière un peu différente
(perception des saveurs).
Les fibres gustatives réagissent à une faible
concentration d'un stimulus gustatif donné, mais répondent
également à des concentrations plus élevées
d'autres goûts (comme dans d'autres sens, par exemple l'audition).
Toutefois, les cellules gustatives présentent des seuils d'excitation
différents.
Les récepteurs de l'amer, détectant en
règle générale les substances toxiques, sont plus sensibles
que les récepteurs de l'acide, eux-mêmes, plus sensibles à
ceux du sucré et du salé.
La transduction des cinq goûts
primaires est différente selon la saveur en cause :
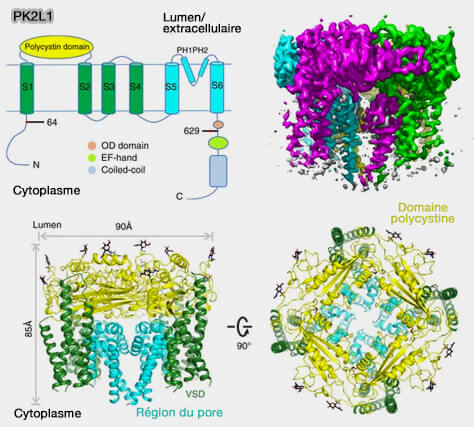
transduction par des cations pour les saveurs salées et acides ;
transduction des molécules sapides par des protéines réceptrices dimériques localisées dans la membrane des cellules réceptrices pour les saveurs amères, sucrées et umami.
Transduction par des cations
Saveur salée
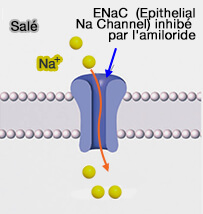
Les ions sodium (Na+) sont principalement à l'origine du goût
salé qui serait perçus par les cellules de type I.
TRPV1 détecte de nombreux sels et serait activé par des concentrations élevées en Na+( rôles de TRPV1)
Lorsque la concentration de Na+ augmente (quand
nous mangeons quelque chose de salé), les ions :
diffusent à
l'intérieur de la cellule gustative (suivant le gradient de Na+),
provoquent un courant qui dépolarise la cellule, ce qui provoque la
libération de neuromédiateurs dans les fibres nerveuses transportant l'information jusqu'au cerveau.
Une telle modulation viendrait compléter les voies centrales de la régulation de l'appétit, sachant que l'ocytocine agit sur les circuits cérébraux pour inhiber la prise de nourriture ( ocytocine et comportement alimentaire).
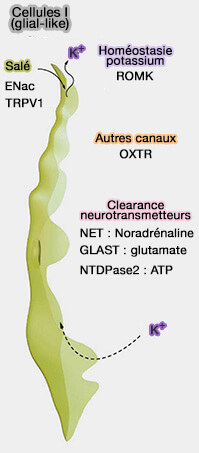
Cellules de type I : autres rôles
Les cellules de type I, outre la perception du goût salé, dégradent et absorbent les neurotransmetteurs comme :
GLAST (GLutamate ASpartate Transporter ou EAAT, Excitatory Amino Acid Transporter) qui est un co-transporteur Na+/glutamate qui intervient dans la saveur umami et peut être VGLUT1, un autre transporteur MFS du glutamate des neurones ;
Les cellules de type I jouent aussi un rôle dans l'homéostasie potassique.
Elles éliminent le K+ extracellulaire qui s’accumule après les potentiels d’action émis par les cellules réceptrices (de type II) et les cellules présynaptiques (de type III).
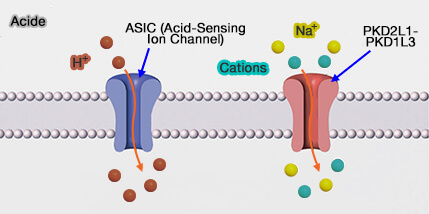
Ce canal n’est pas un canal traditionnel lié au ligand : le stimulus acide ferme le canal et celui-ci ne s'ouvre qu’après le retrait du stimulus : ce mécanisme est appelé " réponse off ". Cette réponse dépolarise la cellule. Ce pourrait être l'hypersalivation lors de perception acide qui pourrait entraîner le décrochage du ligand.
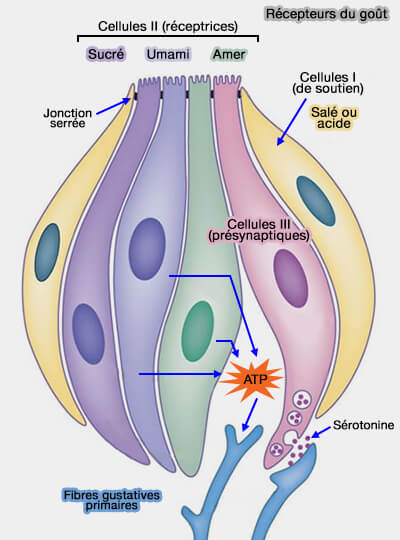
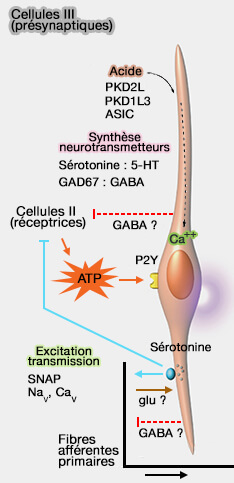
Les cellules présynaptiques de type III peuvent être activées par plusieurs mécanismes distincts :
soit par une variation de pH vue plus haut ;
soit, de façon indirecte, par l’ATP libéré suite à l’activation d’une cellule de type II. Cet ATP se fixe sur des récepteurs purinergiques P2Y4, exprimés à la surface du corps cellulaire. Le récepteur P2Y4 est couplé à une phospholipase C et active la voie de l’IP3 pour permettre la libération du calcium des stocks intracellulaires.
L’augmentation de calcium intracellulaire provoquée par cette activation déclenche la libération de sérotonine par exocytose, et donc l'inhibition de l'ATP.
Qui qu'il en soit, les différentes dépolarisations par ouverture des canaux Na+, K+ ou Ca++ déclenchent le potentiel d'action, en relarguant l'on calcium (Ca++) qui active le complexe SNARE. SNARE permet la fusion des vésicules synaptiques avec la membrane cytoplasmique de la cellule présynaptique (de type III) ce qui provoque :
soit la libération du la noradrénaline ou du GABA pour transporter l'information jusqu'au cerveau ;




 transduction sensorielle).
transduction sensorielle).