transduction des molécules sapides par des protéines réceptrices dimériques localisées dans la membrane des cellules réceptrices pour les saveurs amères, sucrées et umami.
Modèle général de la transduction par des protéines réceptrices dimériques
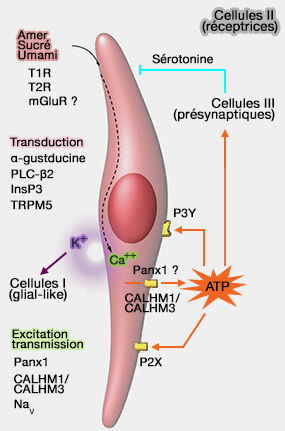
Cellule gustative de type II
(Figure : vetopsy.fr)
Les saveurs amères, sucrées et umami, détectées par les cellules de type II, utilisent les
voies de transduction reliés aux récepteurs couplés aux protéines G (GPCR) .
Les mécanismes moléculaires de cette transduction
sont complexes et mettent en jeu des récepteurs particuliers de la
membrane des cellules gustatives.
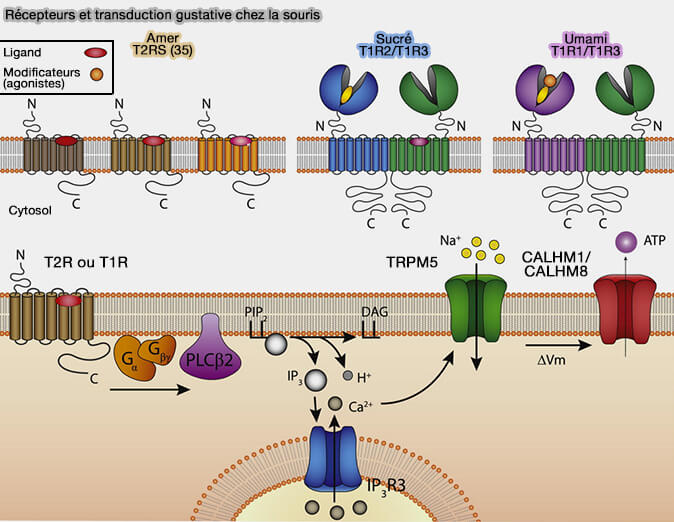
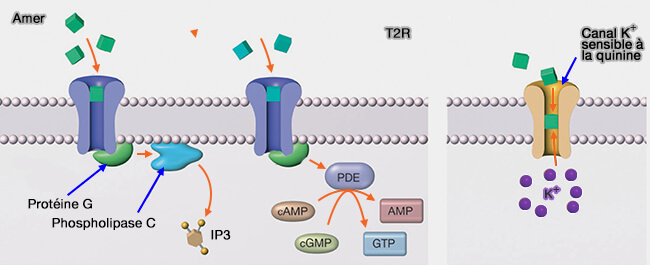
La molécule sapide active son récepteur (T1R ou T2R) couplé à une protéine G (α-gustducine) qui par l’intermédiaire d’une phospholipase C-β2 (PLCB2), produit de l'inositol 1,4,5-trisphosphate (InsP3) et du diacylglycérol (DAG).
En outre, la dépolarisation active la libération d'ATP qui agit à son tour comme neurotransmetteur primaire pour stimuler les récepteurs P2X sur les neurones gustatifs afférents.
Or, les cellules de type II ne possèdent ni vésicules synaptiques, ni expression de gènes intervenant dans leur remplissage.
Récepteurs et transduction gustative (amer, sucré, umami) chez la souris
(Figure : vetopsy.fr d'après Emily L. Liman et coll)
CALHM1, CALcium Homeostasis Modulator 1, canal non sélectif voltage-dépendant, est un élément essentiel à la libération l’ATP en réponse aux potentiels d'action Na+ (CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes 2013).). En son absence, les composés gustatifs ne et Les souris, en l'absence de CALMH1, ne parviennent plus à stimuler la libération d'ATP et perdent la perception des goûts induits par les GPCR (Salty Taste Deficits in CALHM1 Knockout Mice 2014). Cependant, la cinétique d'activation de ces canaux n'est pas satisfaisante.
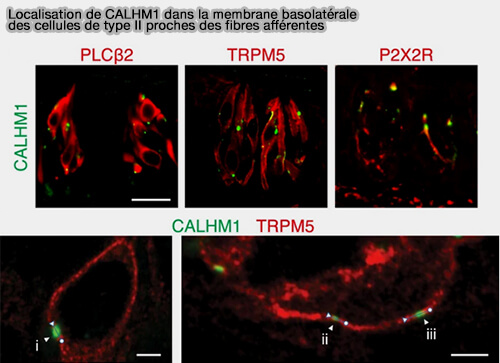
Localisation de CALHM1 dans la membrane basolatérale des
cellules de type II proches des fibres afférentes
(Figure : Kashio et coll)
Calhm3 est coexprimé avec Calhm1 exclusivement dans les les cellules de type II et sa délétion supprime la libération d'ATP provoquée par le goût des papilles gustatives et la perception du goût par le GPCR
indirectement la cellule présynaptique de type III via les récepteurs métabotropiques P2Y4 ou d'autres récepteurs (P2X2, P2X7, P2Y4, P2Y6) présynaptiques. La cellule de type III libère en réponse de la sérotonine (5-HT) qui inhibera en retour la cellule de type II, mettant fin au signal .
Le GABA, libéré également par les cellules de type III, inhiberait la sécrétion d'ATP grâce à la présence de récepteurs GABAA et GABAB sur les cellules réceptrices
L’ADP ou l'adénosine peut exercer un rétrocontrôle positif sur les cellules sensibles au sucré.
Les cellules gustatives expriment de nombreux types de récepteurs sans toutefois les exprimer tous ce qui permet peut-être une certaine distinction (Genetic Labeling of Tas1r1 and Tas2r131 Taste Receptor Cells in Mice 2012). Toutefois, elles sont sensibles à la
plupart des composés amers sans les distinguer.
les " spécialisés " qui détectent un ou plusieurs produits chimiques amers ;
les " généralistes "qui en détectent de nombreux.
Le signal transmis au cerveau est quelque chose comme : « Attention
! Danger ! » et n'a nul besoin d'être spécifique !
Saveurs sucrée et
umami
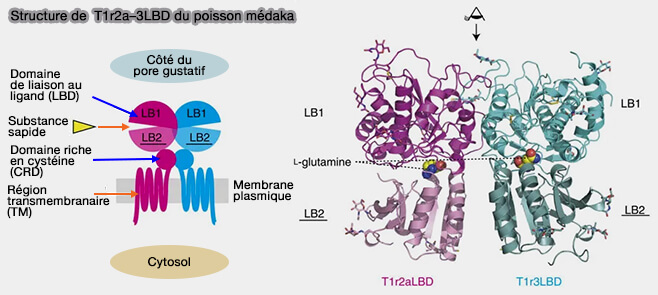
Récepteur T1R
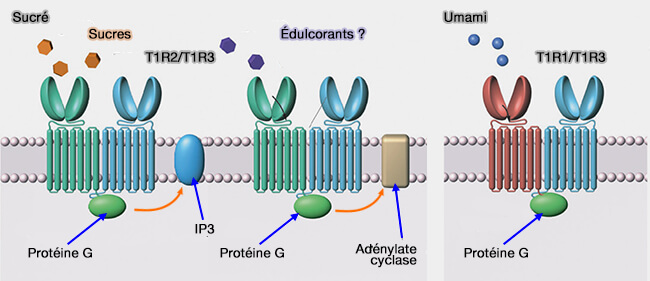
La saveur sucrée et l'umami utilisent les récepteurs T1R (Taste Receptor type 1) qui sont des récepteurs GPCR de classe C qui utilisent l'α-gustducine comme protéine G.
Le sucré, que ce soit pour les sucres naturels et les édulcorants, est perçu par l'hétérodimère T1R2/T1R3;
édulcorants naturels (sucres tels que glucose, fructose, sucrose, maltose), ainsi que certains édulcorants artificiels comme l'aspartame se fixent au domaine extracellulaire de T1R2 ;
édulcorants artificiels, tels que le cyclamate et le lactisole (bloqueur des récepteurs sucrés) ciblent les segments transmembranaires de T1R3,
par le transporteur de glucose de type 4 (GLUT4) ou le co-transporteur sodium/glucose 1 (SGLT1) est dépendant de Na+. On suppose que l'un de ces transporteurs, ou les deux, font partie d'une autre voie de détection du glucose similaire à celle utilisée dans les cellules β du pancréas (Taste buds : cells, signals and synapses 2017).



 généralités sur la transduction gustative) est différente selon la saveur en cause :
généralités sur la transduction gustative) est différente selon la saveur en cause :