Un canal sodium, ou sodique, est un canal ionique, i.e. spécifique aux ions sodium. On en connaît plusieurs catégories selon le mécanisme de passage du Na+ :
1. les canaux sodium dépendants du voltage, voltage-dépendants ou tensiodépendants (" Voltage-gated sodium channel ") dont l'ouverture dépend de la modification de la polarité membranaire ;
2. les canaux sodium dépendants du ligand, ligand-dépendants ou chimiodépendants (" Ligand-gated sodium channel "), classe des récepteurs ionotropes dont les membres s'ouvrent en présence d'un ligand ;
3. les canaux de fuite (nongated channel ou leakage channel) qui laissent passer les ions librement.
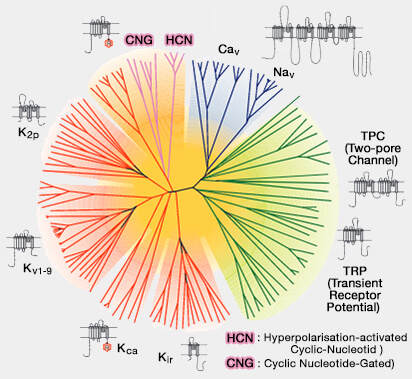
Vue d'ensemble des canaux sodium voltage-dépendants (Nav)
La superfamille des VGL
(Figure : vetopsy.fr d'après Yu et Catterall)
Cependant, contrairement aux autres classes de la superfamille, leurs propriétés fonctionnelles sont relativement similaires.
Ces canaux sont responsables de la phase ascendante rapide du potentiel d'action dans les nerfs, les muscles et les cellules endocrines. Ils sont 10 fois plus rapides que les canaux K+ ou Ca++.
Les médicaments qui bloquent ces canaux sont utilisés pour les anesthésies locales ou celle de la colonne vertébrale, pour le traitement de la douleur, les arythmies cardiaques ou l'épilepsie.
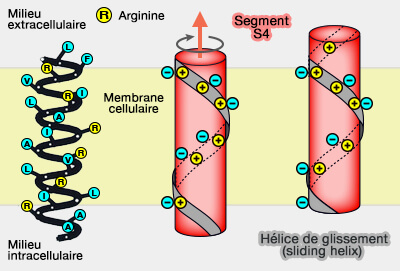
Ces canaux voltage-dépendants s'ouvrent ou se ferment en réponse à une modification de la polarité membranaire.
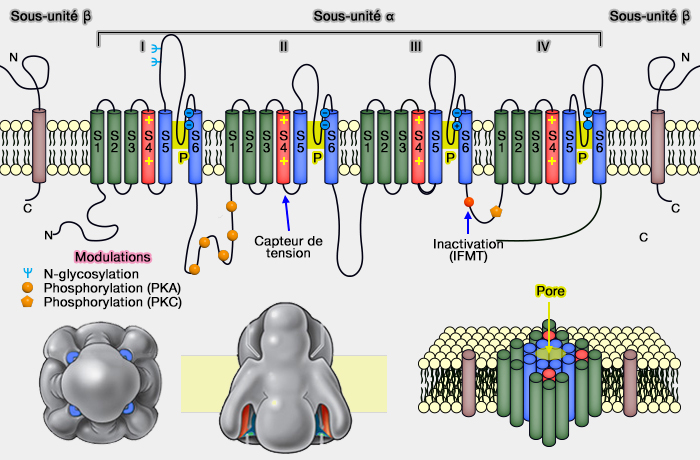
d'une sous-unité α, suffisante pour former le canal Na+ (260 kD) pour le passage des ions,
de sous-unités β qui facilitent leur localisation membranaire et permettent la modulation de leurs propriétés.
Canal Na voltage-dépendant
(Figure : vetopsy.fr)
Sous-unités α
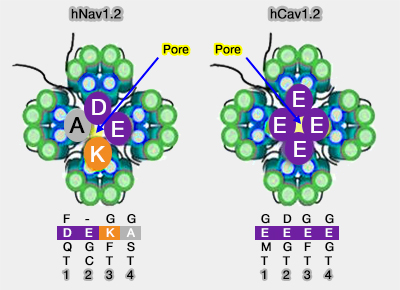
Sélectivité des canaux Na et Ca voltage-dépendants
(Figure : vetopsy.fr d'après Stephens et coll)
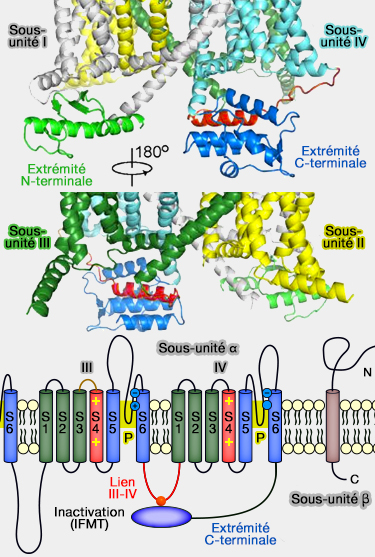
La sous-unité α possède 4 sous-domaines (I à IV), chacun représentant le motif structural commun de la superfamille : cette structure est proche de celle des canaux calcique voltage-dépendants (Cav).
Nav2 / NAG est un canal sodique putatif, dont le rôle physiologique est depuis longtemps une énigme.
La composition des domaines transmembranaire et extracellulaire est homoloque à plus de 50%. Les articles précédents sont très détaillés, mais le tableau de Wikipedia les résume (expression selon les cellules).
On trouve des canaux Nav chez les bactéries (NaChBac - Na-selective Channel of Bacteria) par exemple), identiques aux canaux Kv et formés de 4 sous-unités identiques.
Le motif structural de ces sous-domaines, commun de la superfamille, est composé chacun, comme pour les canaux potassiques voltage dépendants Kv (structure des Kv), de 6 segments transmembranaires (T ou TM) et un pore (P), d'où l'appelation 6TM + 1P.
Les segments S5 et S6 de chaque domaine forment le pore.
La partie du pore proche du milieu extracellulaire comprend deux anneaux sélectifs, dont le filtre sélectif, hautement conservé dans les canaux voltage-dépendants qui joue un rôle différent dans la perméabilité ionique canalaire.
1. L'anneau intérieur est le filtre sélectif DEKA (dit aussi anneau de retrécissement), i.e. aspartate dans le domaine I, glutamate dans le II, lysine dans le III et alanine dans le IV (D372, E898, K1419 et A1711).
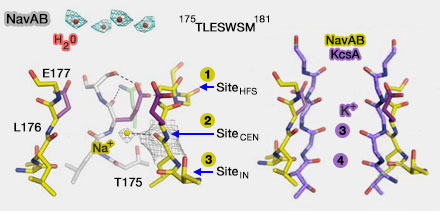
Pour mieux comprendre le fonctionnement des canaux sodiques, deux articles ont on a étudié l'impact de mutations sur les sites du pore de NavAb, un canal sodium voltage-dépendant de la bactérie Arcobacter butzleri. Deux articles sont essentiels pour ces études :
En revanche, le SF des canaux calciques voltage-dépendants Cav est constitué de quatre acides glutamiques, EEEE, tandis que NavAb et NavRh contiennent respectivement TLESWS ou TLSSWE sur chaque protomère.
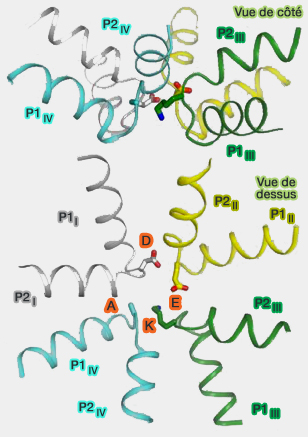
K du motif DEKA et sa chaine
(Figure : vetopsy.fr d'après Ahmed et coll)
Le rôle de la lysine (K) est primordial dans sa sélectivité au sodium, malgré un diamètre presque identique à l'ion calcique.
Le remplacement par une alanine abolit complètement la sélectivité du canal.
La mutation des résidus DEKA en glutamates (EEEE) est sélectif du Ca++.
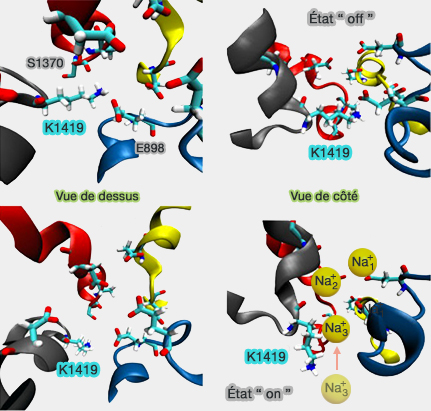
l'état " off " l'en empêche. Dans cet état, la chaîne latérale de K1419 interagit avec le groupe carboxylate de E898 du filtre DEKA et les atomes d'oxygène de la chaîne latérale de S1710, un résidu de la paroi des pores residues, ce qui lui permet de bloquer le pore pour tous les cations (Conformational changes of the pore domain segments between NavPaS and Cav1.1 - 2017 -).
Ce processus pourrait également bloquer le déplacement en sens inverse de l'ion lors de l'activation.
L'étroitesse du pore de 0,3 à 0,5 nm, ne permet que le passage d'un ion Na+ et d'une molécule d'eau, les cations plus gros, comme le K+, ou des acides aminés positifs ne peuvent s'y engouffrer.
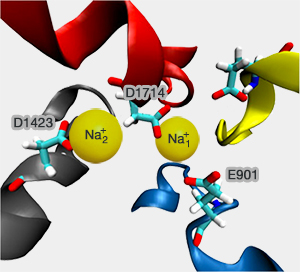
Motif EEDD de l'anneau extérieur
(Figure : vetopsy.fr d'après Ahmed)
2. L'anneau extérieur EEDD est formé de quatre acides aminés, i.e. glutamate dans le domaine I et II, aspartate dans le III et le IV (E375, E901, D1423 et D1714) qui garde l'entrée du vestibule.
Porte d'inactivation
(Figure : vetopsy.fr d'après Shen et coll)
Cette inactivation exerce un contrôle sur la conductance du canal de sodium, i.e. capacité d'un corps, soumis à une différence de potentiel, à laisser passer une certaine quantité de courant électrique, ce qui permet une succession rapide d'ouvertures et de fermetures du canal pour générer des salves rapides et brefs de potentiel d'action.
La présence d'une courte hélice après le motif IFMTF
participerait au phénomène en participant à une interaction avec la terminaison COOH du canal pour stabiliser la fermeture de la porte et minimiser la réouverture de canal (The Na+ Channel Inactivation Gate Is a Molecular Complex 2004).
D'autres inactivations plus lentes sont possibles et ne font pas intervenir cette porte, mais sont dépendantes de la conformation du filtre séléctif.
Autres domaines
1. Les domaines extracellulaires sont représentés par :
Ψ, les sites probables de N-glycosylation dans le domaine I sur le lien S5-S6 ;
Pour les canaux voltage-dépendant, si les sous-unités α sont impliquées dans la conductance ionique, des sous-unités complémentaires en modulent les multiples aspects de leur comportement et jouent un rôle crucial dans le contrôle de l'excitabilité neuronale.





 expression selon les cellules).
expression selon les cellules). 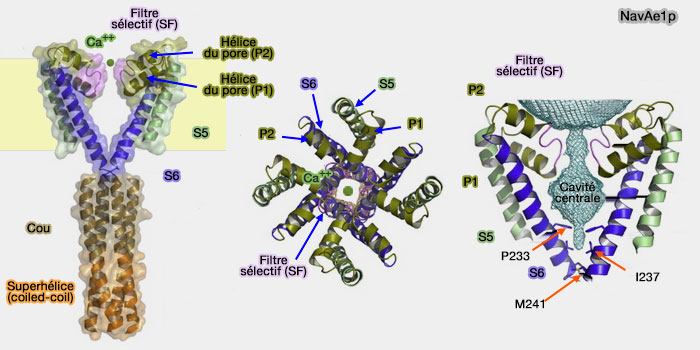
 Conformational changes of the pore domain segments between NavPaS and Cav1.1 - 2017 -).
Conformational changes of the pore domain segments between NavPaS and Cav1.1 - 2017 -).