La membrane plasmique est une bicouche lipidique asymétrique organisée en domaines dynamiques où lipides et protéines assurent cohésion structurale et compartimentation cellulaire.
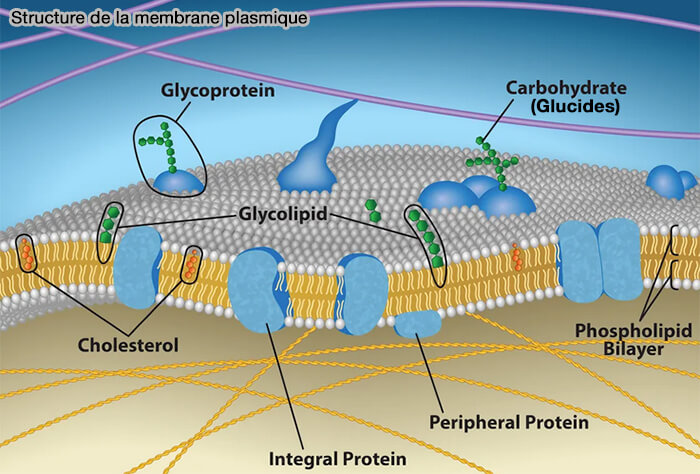
Structure de la membrane plasmique
La membrane plasmique est constituée d’une bicouche lipidique amphiphile formant une barrière physico-chimique continue entre le cytoplasme et le milieu extracellulaire.
Structure de la membrane plasmique
(Figure : vetopsy.fr d'après openstax)
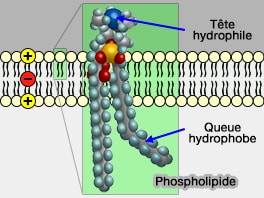
Organisation de la bicouche lipidique
1. Les phospholipides s’organisent en double feuillet, avec les têtes hydrophiles orientées vers les milieux aqueux et les chaînes hydrophobes regroupées au centre de la membrane.
Phospholipide membranaire
(Figure : vetopsy.fr)
Cette organisation génère une structure d’environ 4-5 nm d’épaisseur assurant une barrière sélective aux molécules polaires ou chargées et constitue l’architecture fondamentale commune à l’ensemble des membranes biologiques.
2. La bicouche lipidique présente une composition complexe déterminant ainsi ses propriétés physiques, notamment sa fluidité, sa perméabilité, sa courbure et son organisation latérale.
Elle est dominée par :
les phospholipides, qui constituent la matrice structurale,
le cholestérol, qui module la fluidité, la rigidité et la stabilité mécanique de la bicouche
les glycolipides, formés par un lipide lié à un ose au sens large par une liaison glycosidique, sont majoritairement localisés dans le feuillet externe de la membrane plasmique et participent aux interactions cellulaires et aux mécanismes de reconnaissance.
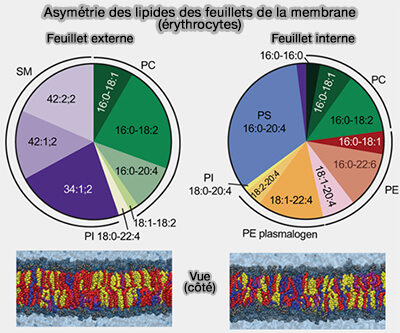
Asymétrie des lipides des feuillets membranaires
(Figure : vetopsy.fr d'après Lorent et coll)
3. Les deux feuillets de la membrane plasmique présentent une composition lipidique différente.
a. Elle confère à la bicouche une asymétrie structurale et fonctionnelle.
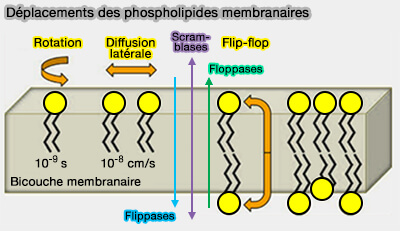
b. Cette asymétrie est maintenue par l’action coordonnée de transporteurs lipidiques spécifiques, notamment les flippases, floppases et scramblases, et joue un rôle essentiel dans :
la signalisation cellulaire,
la polarité membranaire,
Déplacement des phospholipides membranaires
(Figure : vetopsy.fr d'après Pomorski et coll)
les processus de reconnaissance.
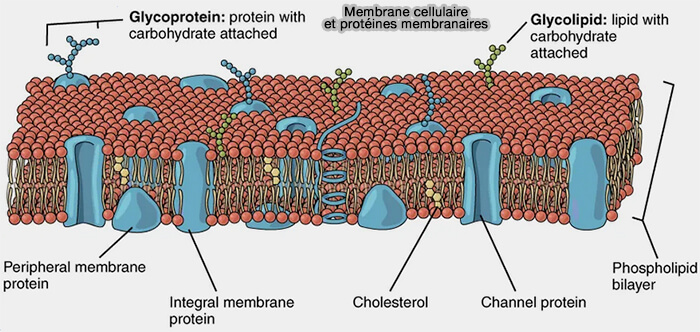
Protéines membranaires
La membrane plasmique contient des protéines associées à la bicouche lipidique selon des modes d’interaction distincts, définissant des catégories structurales qui ne se recouvrent pas nécessairement.
des protéines monotopiques, insérées dans un seul feuillet membranaire sans traverser la bicouche,
des protéines ancrées aux lipides, associées à la membrane par une modification lipidique covalente (ancre GPI, acylation, prénylation) sans segment transmembranaire.
La répartition de ces différentes catégories de protéines conditionne les fonctions de transport, de signalisation et d’organisation structurale des membranes biologiques.
Membrane cellulaire et protéines membranaires
(Figure : vetopsy.fr d'après openstax)
Propriétés physiques de la bicouche lipidique
La bicouche lipidique possède des propriétés mécaniques et physico-chimiques qui résultent directement de sa composition moléculaire.
1. Sa perméabilité intrinsèque dépend de la région centrale hydrophobe et de l’organisation amphiphile des phospholipides qui constitue une barrière énergétique ( transports membranaires) :
permettant le passage des petites molécules apolaires,
limitant le passage spontané des ions et des molécules polaires.
2. L’épaisseur moyenne de 4-5 nm est déterminée par la longueur des chaînes acyles (R-C(=O)-).
Les segments transmembranaires des protéines doivent présenter une longueur hydrophobe compatible avec cette épaisseur, conditionnant leur insertion stable dans la membrane.
3. La courbure dépend de la géométrie des lipides constituants (courbures membranaires).
Les lipides de forme cylindrique favorisent des surfaces planes, tandis que les lipides de forme conique ou inversée (comme la phosphatidyléthanolamine) favorisent des déformations locales et des courbures négatives.
L’asymétrie lipidique entre les feuillets contribue également aux contraintes de courbure.
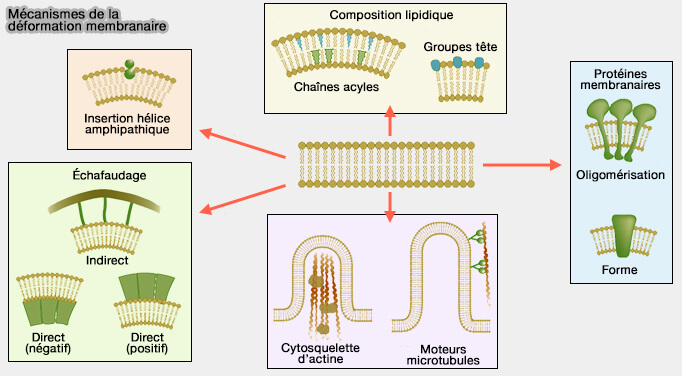
Mécanisme de déformation membranaire
(Figure : vetopsy.fr d'après Mc Mahon et coll)
4. La membrane plasmique présente une élasticité limitée lui permettant de se déformer transitoirement sous contrainte mécanique.
Ces propriétés sont modulées par la composition lipidique et par le couplage au cytosquelette cortical.
Évolution des modèles conceptuels de la membrane plasmique
L’organisation de la membrane plasmique a fait l’objet d’une évolution conceptuelle progressive, depuis les premières représentations structurales jusqu’aux modèles intégrant hétérogénéité latérale et couplage cytosquelettique (Membrane models).
Modèle fondateur de l’organisation membranaire
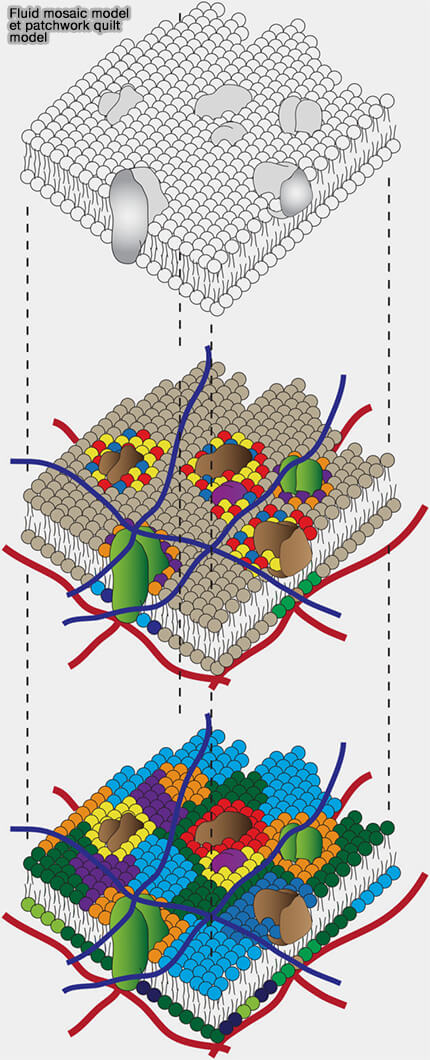
Le modèle de la mosaïque fluide (fluid mosaic model), proposé par Singer et Nicolson en 1972, établit que la membrane plasmique correspond à une bicouche lipidique fluide au sein de laquelle les protéines diffusent latéralement dans un environnement dynamique (The Fluid Mosaic Model of the Structure of Cell Membranes 1972).
Les lipides assurent la cohésion structurale, tandis que les protéines remplissent des fonctions spécifiques sans former une couche rigide continue.
1. La diffusion latérale des phospholipides est rapide et homogène à l’échelle moléculaire.
3. Ce modèle considère principalement la membrane comme une entité autonome.
Cependant, la membrane plasmique est en réalité insérée dans un environnement structural plus large en établissant des interactions avec la matrice extracellulaire du côté externe et avec le cytosquelette cortical du côté interne.
Ces interactions mécaniques et fonctionnelles n’étaient pas intégrées dans la formulation initiale du modèle.
Modèles contemporains
Les développements expérimentaux postérieurs au modèle de la mosaïque fluide ont conduit à réévaluer l’organisation membranaire en mettant en évidence une hétérogénéité latérale et un couplage étroit avec le cytosquelette cortical.
1. Le modèle " patchwork " (patchwork quilt model) introduit une organisation latérale hétérogène, dans laquelle certaines régions membranaires présentent une composition lipidique et protéique distincte, formant des domaines dynamiques.
Ces représentations ne s’excluent pas mutuellement : la fluidité latérale demeure une propriété fondamentale, mais elle s’inscrit dans une organisation plus structurée et spatialement modulée.
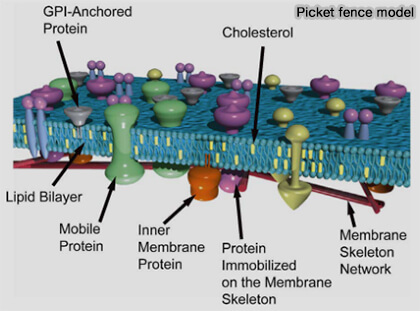
Picket fence model
(Figure vetopsy.fr d'après Ritchie et coll)
Dans cette perspective limitant la diffusion latérale et induisant une diffusion par sauts entre compartiments :
certaines protéines transmembranaires ancrées au cytosquelette jouent le rôle de " piquets ",
le réseau d’actine forme une " clôture ".
3. Un autre modèle " active actin-membrane composite model " décrit la membrane comme un système mécaniquement couplé au cytosquelette cortical et influencé par des processus actifs dépendants de l’ATP.
Cette approche met en avant le rôle de la polymérisation et de la dépolymérisation dynamiques de l’actine, de l’activité des moteurs moléculaires tels que les myosines et des forces mécaniques générées au cortex cellulaire dans le remodelage continu de l’organisation latérale.
Les flux d’énergie dépendants de l’ATP permettent des réarrangements transitoires du réseau cortical, modifiant localement la tension membranaire, la distribution des protéines et la stabilité des nanodomaines.
La membrane n’est ainsi plus considérée comme une surface simplement compartimentée, mais comme un système mécaniquement actif dont l’organisation résulte d’interactions dynamiques entre bicouche lipidique, cytosquelette et contraintes physiques.
Cette évolution conceptuelle ne remplace pas le modèle de la mosaïque fluide, mais le complète en intégrant la dimension structurale, mécanique et dynamique de l’organisation membranaire.
Domaines membranaires et hétérogénéité structurale
L’organisation latérale influence directement la répartition des récepteurs, la formation de plateformes de signalisation et la coordination des interactions protéine-protéine au sein de la membrane.
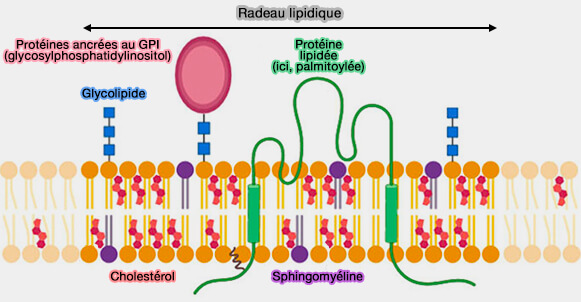
1. Les membranes biologiques contiennent des microdomaines spécialisés, souvent désignés sous le terme de radeaux lipidiques, caractérisés par un enrichissement en cholestérol et en sphingolipides.
a. Ces domaines présentent une organisation plus ordonnée que le reste de la bicouche et constituent des régions de composition lipidique particulière en concentrant préférentiellement certaines protéines membranaires, notamment :
Structure des radeaux lipidiques
(Figure : vetopsy.fr d'après Ripa et coll)
b. Les radeaux lipidiques sont dynamiques et réversibles.
Ils ne correspondent pas à une catégorie structurale distincte de protéines membranaires, mais à une organisation lipidique spécifique modulant leur distribution et leur activité fonctionnelle.
2. La diffusion latérale des protéines membranaires n’est pas libre sur toute la surface cellulaire.
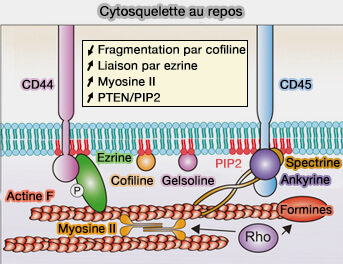
Cytosquelette au repos
(Figure : vetopsy.fr d'après Ostrowski et coll)
a. Elle est restreinte par l’organisation du cytosquelette cortical, notamment par le réseau d’actine situé sous la membrane plasmique.
Ce réseau forme une trame sous-membranaire à laquelle sont reliées certaines protéines transmembranaires par l’intermédiaire de protéines d’ancrage, comme les adaptateurs.
Cette organisation crée des compartiments membranaires de petite taille dans lesquels les protéines diffusent de manière limitée avant de franchir des barrières cytosquelettiques.
b. Ce mécanisme, parfois décrit comme un modèle de " clôture et piquets " (picket fence model), explique que la membrane plasmique soit structurée en domaines dynamiques définis par l’interaction entre bicouche lipidique et cytosquelette (Toward a new picture of the living plasma membrane 2020).
a. Dans ce cadre, l’organisation latérale ne résulte pas uniquement de propriétés biophysiques globales de la bicouche ou de contraintes imposées par le cytosquelette, mais d’une combinatoire spécifique entre lipides et protéines.
Certaines protéines reconnaissent des environnements lipidiques particuliers, tandis que certaines compositions lipidiques stabilisent préférentiellement des ensembles protéiques définis.
L’organisation membranaire émerge ainsi d’interactions coopératives lipide-protéine qui déterminent la localisation, la dynamique et la fonction des complexes membranaires.
b. Cette proposition ne remplace pas les modèles conceptuels antérieurs ( modèles membranaires), mais introduit un niveau supplémentaire d’interprétation en suggérant que les zones fonctionnelles de la membrane correspondent à des signatures moléculaires spécifiques combinant lipides et protéines.
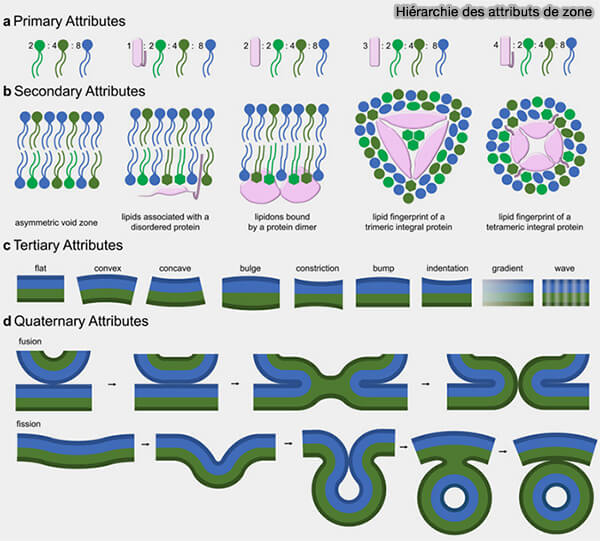
Hiérarchie des attributs de zone sans le modèle proteolipid code
(Figure : vetopsy.fr d'après Kerwin et Overduin)
Remarque : dans la figure, les nombres associés aux protéines (1 à 4) indiquent le nombre d’espèces protéiques différentes regroupées dans une zone donnée, tandis que les nombres associés aux lipides (2, 4, 8) correspondent au nombre de types lipidiques distincts présents dans l’environnement local, représentés par des couleurs différentes.
L’augmentation conjointe de la diversité protéique et lipidique illustre le principe du « proteolipid code » : la fonction d’un domaine membranaire dépend de la combinaison spécifique entre un ensemble protéique et une signature lipidique locale.





 transports membranaires) :
transports membranaires) :