La condensation et la décondensation chromosomiques assurent la transformation réversible de la chromatine en chromosomes mitotiques fonctionnels, conditionnant la ségrégation fidèle du génome.
Vue d'ensemble
1. La condensation chromosomique constitue un événement structural majeur de l’entrée en mitose.
Elle transforme la chromatine interphasique diffuse en chromosomes individualisés, compacts et mécaniquement manipulables, adaptés à leur prise en charge par le fuseau mitotique.
Ce remodelage structural assure trois fonctions essentielles :
2. La décondensation chromosomique constitue le phénomène inverse, strictement contrôlé dans le temps.
Elle débute en télophase après la séparation irréversible des chromatides et permet la restauration d’une chromatine interphasique fonctionnelle, compatible avec la reprise des activités nucléaires.
Déclenchement de la condensation
La condensation chromosomique s’engage dès l’entrée en mitose, sous l’effet de la réorganisation globale des équilibres enzymatiques qui caractérise la transition G2/M.
L’activation rapide du complexe cycline B/CDK1 (MPF) impose un régime de phosphorylation mitotique généralisée, relayé par Plk1 et Aurora B, qui modifie profondément les propriétés de la chromatine et de ses régulateurs structuraux.
1. La phosphorylation des histones, notamment H1 et H3, par CDK1 et Aurora B
diminue l’affinité de l’histone H1 pour l’ADN,
induit la phosphorylation de H3 sur la sérine 10 (H3S10ph).
La nouvelle organisation des nucléosomes et les interactions entre fibres de chromatine favorisent alors le passage d’une chromatine interphasique souple vers une structure plus rigide, plus segmentée et plus propice à la compaction.
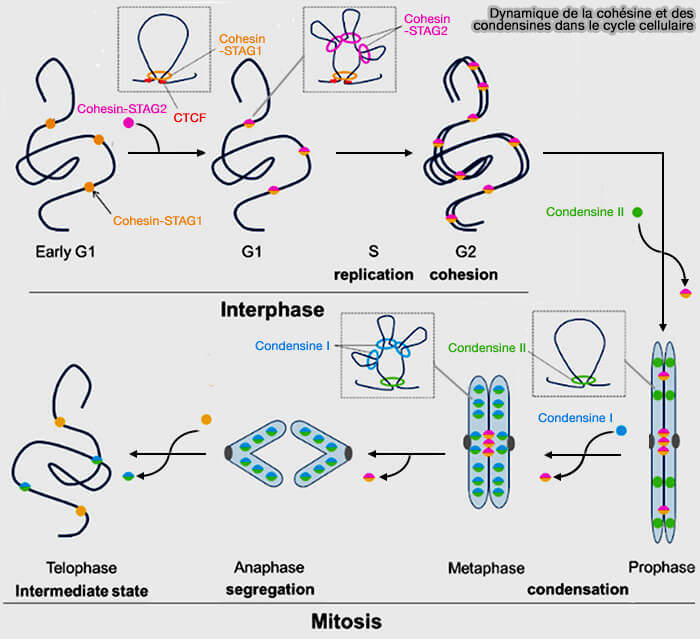
Dynamique de la cohésine et des condensines dans le cycle cellulaire
Figure : vetopsy.fr d'après Ryzhkova et coll)
2. La désorganisation de l’architecture nucléaire accompagne ce processus.
Les condensines sont activées par phosphorylation mitotique (CDK1, Plk1) et s’accumulent progressivement sur la chromatine, où elles organisent l’ADN par extrusion de boucles et structuration autour d’un axe protéique central ( accumulation des condensines).
La condensine II agit dès la prophase pour établir l’architecture longitudinale des chromatides, tandis que la condensine I, recrutée après rupture de l’enveloppe nucléaire, renforce la compaction en resserrant les boucles et en stabilisant la structure métaphasique.
Les condensines sont étudiées dans un chapitre spécifique.
b. En parallèle, les cohésines sont progressivement éliminées des bras chromosomiques sous l’effet de la phosphorylation mitotique et de l’ouverture contrôlée de l’anneau par WAPL, ce qui permet l’individualisation morphologique des chromatides ( voie prophasique de dissociation).
La cohésion centromérique est en revanche protégée par le complexe Sgo1-PP2A, assurant le maintien de l’association des chromatides sœurs jusqu’à l’anaphase et permettant l’établissement de la tension métaphasique ( protection centromérique).
3. L’ensemble de ces événements conduit à l’apparition progressive de chromosomes visibles, compacts et mécaniquement cohérents, capables d’être pris en charge par le fuseau mitotique en prométaphase.
Architecture fonctionnelle du chromosome mitotique
Le chromosome mitotique est une structure transitoire spécialisée, optimisée pour transmettre les forces du fuseau mitotique et assurer la ségrégation fidèle des chromatides.
1. Chaque chromosome apparaît comme une entité individualisée constituée de deux chromatides sœurs parallèles, reliées par une région centromérique mécaniquement résistante.
Cette organisation repose sur plusieurs caractéristiques morphologiques stables en métaphase :
organisation hélicoïdale globale contribuant à la compacité,
diamètre relativement homogène des chromatides,
individualisation nette des chromosomes les uns par rapport aux autres.
2. La cohésion entre chromatides sœurs persiste sélectivement au niveau centromérique, ce qui confère aux chromosomes mitotiques leurs propriétés fonctionnelles en métaphase.
3. Les chromosomes mitotiques possèdent ainsi des propriétés mécaniques spécifiques adaptées à leur fonction :
rigidité suffisante pour transmettre les forces exercées par les microtubules kinétochoriens sans déformation excessive de l’axe chromosomique,
élasticité contrôlée permettant d’absorber les fluctuations mécaniques du fuseau.
Ces propriétés rendent possible la biorientation, la stabilité en métaphase et la séparation fidèle des chromatides.
Remarque : la condensation et la décondensation chromosomiques doivent être étroitement coordonnées avec la dynamique du fuseau et le cycle mitotique car toute altération de ces processus compromet la fidélité de la ségrégation.
Une condensation insuffisante favorise des défauts d’attachement kinétochorien, des erreurs de biorientation et des retards de ségrégation, conduisant à des chromosomes traînards, i.e. des chromosomes qui migrent mal en anaphase, pouvant être exclus du noyau, former des micronoyaux et favoriser l’instabilité génomique et l’aneuploïdie.
À l’inverse, une décondensation incomplète perturbe la reconstitution correcte du noyau en télophase, avec des conséquences possibles sur l’organisation chromatinienne, l’expression génique et la stabilité du génome.
Décondensation chromosomique
1. La décondensation chromosomique correspond au retour progressif des chromosomes mitotiques vers une chromatine interphasique fonctionnelle après la séparation irréversible des chromatides.
Elle ne constitue pas un simple relâchement passif, mais un processus strictement coordonné avec la sortie de mitose, initié en anaphase B sur le plan moléculaire et devenant morphologiquement visible en télophase.
a. Elle est dépendante de l’extinction de l’activité du complexe cycline B/CDK1 (MPF) par l’ubiquitination de la cycline B par l’APC/CCdc20, ce qui entraîne l’extinction globale de la phosphorylation mitotique.
b. Les phosphatases PP1 et PP2A deviennent prépondérantes et assurent la déphosphorylation coordonnée de nombreux substrats mitotiques :
des histones H1 et H3, favorisant le relâchement de la compaction,
des condensines et de la topoisomérase IIα, contribuant à la perte progressive de la rigidité chromosomique,





 accumulation des condensines).
accumulation des condensines).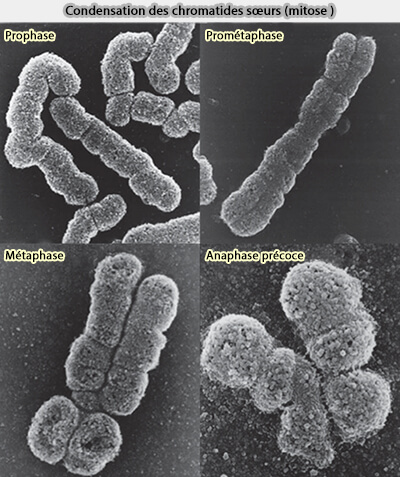
{kind=link}