Les condensines structurent les chromosomes mitotiques par extrusion de boucles d’ADN, générant la compaction, la rigidité et l’individualisation des chromatides.
Les condensines : organisation chromosomique
Définition et positionnement fonctionnel
Les condensines sont des complexes SMC spécialisés dans l’organisation tridimensionnelle des chromosomes mitotiques.
1. Elles ne participent pas à la cohésion, mais assurent :
la compaction longitudinale des chromatides,
la rigidification mécanique des axes chromosomiques,
la structuration en domaines et en hélices régulières,
la résolution des contraintes topologiques nécessaires à une ségrégation fidèle
2. Leur fonction est fondamentalement architecturale : elles traduisent l’activité enzymatique des complexes SMC en propriétés mécaniques mesurables du chromosome mitotique (compaction, rigidité, individualisation).
3. Les cellules eucaryotes possèdent deux complexes distincts, fonctionnellement complémentaires.
La condensine I, qui intervient principalement dans la compaction latérale et l’organisation fine des boucles, générant une forte densité de bouclage en métaphase et conférant aux chromosomes leur compacité maximale.
La condensine II intervient précocement pour structurer l’axe chromosomique, assurant l’allongement et l’individualisation précoce des chromatides et déterminant l’architecture globale du chromosome mitotique.
Ces deux complexes coopèrent de manière hiérarchique selon des cinétiques distinctes.
Composition moléculaire du complexe condensine
Les deux complexes condensine correspondent à une déclinaison fonctionnelle spécialisée de l’architecture des complexes SMC.
1. Le cœur du complexe est constitué d’un hétérodimère formé par SMC2 et SMC4 qui permet la formation d’une structure en anneau capable d’encercler topologiquement l’ADN, base mécanique de l’extrusion de boucles et de l’organisation chromosomique.
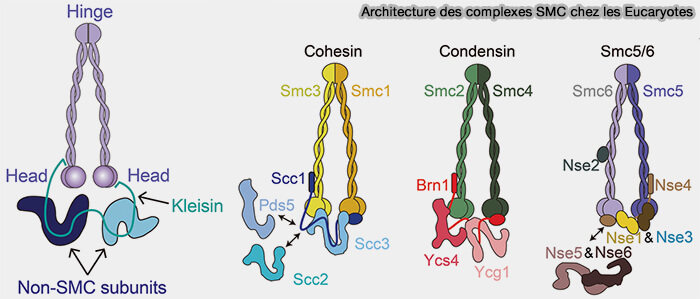
Architecture des complexes SMC des Eucaryotes
(Figure : vetopsy.fr d'après Moon et Ryu)
2. Les propriétés spécifiques du complexe découlent de l’association du dimère SMC2-SMC4 avec des sous-unités non-SMC spécialisées.
a. La kleisine relie directement les deux têtes ATPases de SMC2 et SMC4 et ferme l’anneau condensine.
CAP-H2 est la kleisine de la condensine II.
CAP-H est la kleisine de la condensine I.
b. Les sous-unités riches en répétitions HEAT contribuent à la stabilisation du complexe sur la chromatine et à la formation d’une plateforme d’interaction pour les régulateurs des condensines.
CAP-D3 et CAP-G2 confèrent à la condensine II une capacité accrue à structurer l’axe chromosomique profond.
CAP-D2 et CAP-G de la condensine I favorisent un bouclage plus dense et plus périphérique.
Organisation fonctionnelle des condensines I et II
La condensine I est plus dynamique, avec un turnover plus rapide.
2. Les deux complexes agissent à des échelles distinctes et selon une organisation spatiale différenciée.
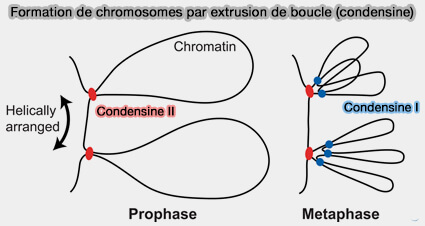
a. La condensine II génère préférentiellement des boucles de grande taille, peu nombreuses, qui structurent l’architecture globale et l’axe longitudinal des chromosomes.
Elle est enrichie vers les régions axiales internes et contribue à la cohérence profonde du chromosome mitotique.
b. La condensine I génère préférentiellement des boucles plus courtes, beaucoup plus nombreuses, responsables de la compaction fine et de la densité élevée de bouclage observée en métaphase.
Elle se distribue de manière plus périphérique et dynamique le long des chromatides.
Condensines et cycle cellulaire
Contrairement aux cohésines, le fonctionnement des condensines ne repose pas sur un mécanisme de chargement dédié équivalent au complexe NIPBL-MAU2.
La compartimentation subcellulaire constitue un déterminant majeur de l’accessibilité des condensines aux chromosomes.
L’activation par phosphorylation à l’entrée en mitose conditionne l’acquisition d’une affinité stable pour la chromatine et le déclenchement de l’activité architecturale.
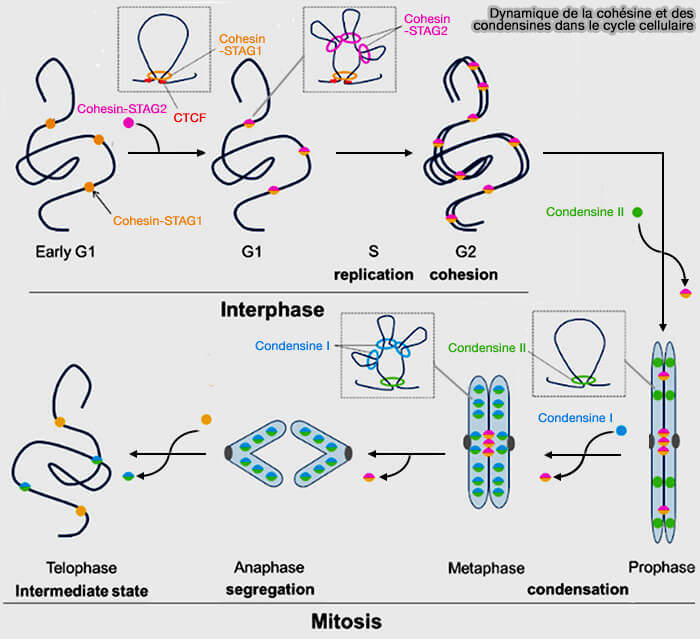
Dynamique de la cohésine et des condensines dans le cycle cellulaire
Figure : vetopsy.fr d'après Ryzhkova et coll)
Interphase
En interphase, les condensines sont présentes mais restent globalement inactives sur le plan architectural, sans effet morphologique détectable au niveau cytologique.
La condensine II est localisée dans le noyau, tandis que la condensine I est majoritairement cytoplasmique, en raison de l’intégrité de l’enveloppe nucléaire.
L’organisation de la chromatine reste typiquement interphasique, dominée par les compartiments A/B (séparation fonctionnelle euchromatine/hétérochromatine) et par les TAD (Topologically Associating Domain), qui correspondent à des domaines d’interactions locales stabilisés principalement par l’activité des cohésines.
L’absence d’activité du complexe cycline B/CDK1 (MPF) et la faible phosphorylation des sous-unités régulatrices maintiennent les condensines dans un état faiblement associé à la chromatine.
Dans certains contextes cellulaires, la condensine II peut néanmoins contribuer à la stabilisation de domaines chromatiniens spécifiques, notamment dans l’hétérochromatine, suggérant un rôle architectural mineur mais réel en dehors de la mitose.
Prophase
En prophase, l’activation du complexe cycline B/CDK1 (MPF) induit la phosphorylation de plusieurs sous-unités régulatrices des condensines, en particulier les kleisines (CAP-H2 pour la condensine II) et les sous-unités HEAT (CAP-D3, CAP-G2), déclenchant leur activation fonctionnelle et leur recrutement stable sur la chromatine.
Elle est ensuite modulée par d’autres kinases mitotiques.
Plk1 contribue au renforcement du recrutement chromatinien.
Aurora B participe à une régulation spatiale locale, notamment au niveau centromérique.
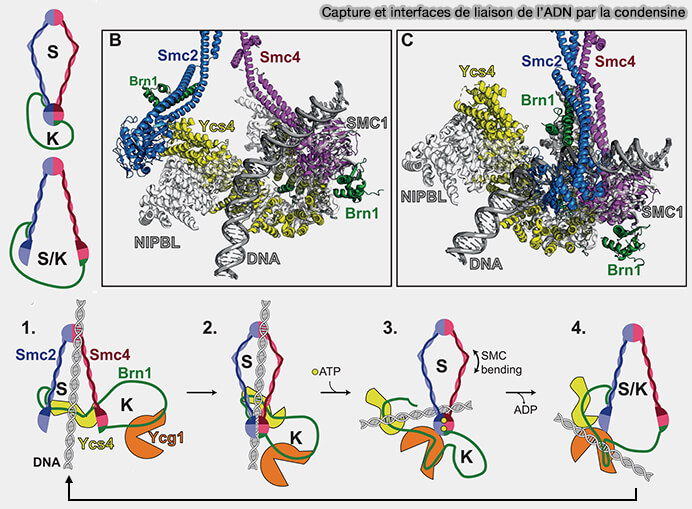
Capture et interfaces de liaison de l’ADN par la condensine
Figure : vetopsy.fr d'après Cutts et Vanini)
1. La condensine II devient alors fonctionnellement active et commence à s’accumuler sur la chromatine, ce qui se traduit par des effets structuraux précoces et encore limités, directement liés aux propriétés mécaniques et cinétiques de la condensine II à ce stade.
La mise en place d’une organisation axiale longitudinale résulte de l’extrusion de grandes boucles par la condensine II, dont l’alignement des points d’ancrage fait émerger un axe chromosomique central.
La cohérence longitudinale initiale des chromatides est amorcée par les cohésines, puis transformée en un axe chromosomique mitotique rigidifié par l’action de la condensine II.
La condensation reste modérée à ce stade, car la densité de complexes actifs sur la chromatine demeure limitée, ce qui restreint le degré de repliement hiérarchique des boucles et maintient les chromosomes visibles mais encore allongés.
La rigidité mécanique demeure faible à intermédiaire, reflétant un maillage structural encore insuffisant pour conférer la cohérence mécanique caractéristique des chromosomes métaphasiques.
2. L’architecture hélicoïdale caractéristique des chromosomes mitotiques n’est pas encore pleinement établie à ce stade.
Prométaphase
La prométaphase correspond à une phase de transition structurale majeure, au cours de laquelle l’architecture chromosomique passe d’une organisation axiale initiale à une organisation mitotique quasi mature.
En prométaphase, l’activité de Plk1 devient un déterminant majeur de la stabilisation de la condensine I sur les chromosomes, contribuant à l’augmentation de la densité de complexes actifs et à la maturation rapide de l’architecture mitotique.
1. La rupture de l’enveloppe nucléaire permet l’accès massif de la condensine I aux chromosomes, qui étaient jusque-là exclusivement structurés par la condensine II.
L’arrivée de la condensine I entraîne une augmentation brutale de la densité globale de complexes condensines actifs sur la chromatine.
2. Cette augmentation de densité modifie profondément la dynamique d’organisation chromosomique.
a. L’amplification de l’extrusion de boucles résulte à la fois du nombre accru de complexes actifs et des propriétés intrinsèques de la condensine I, qui favorise la formation de boucles plus courtes et plus nombreuses.
Cette multiplication des points d’ancrage structuraux induit un repliement plus serré de la chromatine et une augmentation rapide de la compaction.
b. La mise en place progressive d’une organisation hélicoïdale régulière des chromatides découle de la superposition hiérarchique des boucles générées par la condensine II (organisation globale) et par la condensine I (organisation fine), conduisant à une architecture tridimensionnelle hautement ordonnée.
L’alignement des points d’ancrage des boucles le long d’un axe central impose des contraintes spatiales entre boucles voisines et transforme la chromatine en un polymère fortement contraint.
Dans ce contexte, la géométrie hélicoïdale correspond à l’état mécaniquement le plus stable en permettant de répartir les contraintes dans l’espace, en limitant les interférences entre boucles et en optimisant la compaction tout en conservant la cohérence structurelle des chromatides.
c. L’augmentation nette de la rigidité mécanique des chromatides résulte de la densification du réseau de boucles imposé par les condensines, qui transforme la chromatine d’un polymère flexible en une structure mécaniquement intégrée.
Ce maillage contraint limite les déformations locales, répartit les forces le long de l’axe chromosomique et confère aux chromatides la résistance nécessaire aux forces de traction exercées par le fuseau mitotique.
Métaphase
En métaphase, l’architecture mitotique est pleinement établie et la géométrie globale du chromosome devient stable.
L’activité des condensines I et II atteint un plateau fonctionnel, soutenu par une phosphorylation mitotique élevée et une densité de fixation homogène le long des chromatides.
Les condensines maintiennent ainsi un état structural des chromosomes indispensable à leur alignement métaphasique et à leur résistance aux forces de traction exercées par le fuseau.
Anaphase
Vue d'ensemble
En anaphase, la chute d’activité du complexe cycline B/CDK1 (MPF) entraîne une déphosphorylation progressive des condensines, sans provoquer leur dissociation brutale de la chromatine.
Les deux complexes restent majoritairement associés aux chromatides pendant leur migration vers les pôles, mais leur dynamique devient progressivement moins stabilisée.
La structure hélicoïdale globale persiste pendant l’essentiel de l’anaphase, puis commence à s’atténuer progressivement à mesure que l’état de phosphorylation mitotique décroît.
Sur le plan fonctionnel, les condensines assurent à ce stade un rôle principalement mécanique et topologique.
Le maintien de la compaction et la résistance à la traction du fuseau résultent de la persistance du réseau de boucles établi en métaphase, qui confère aux chromatides une cohérence structurale suffisante pour éviter leur étirement excessif.
le maintien transitoire de l’organisation imposée par les condensines,
l’action coordonnée de la topoisomérase IIα
1. Le maintien transitoire de l’organisation imposée par les condensines joue un rôle essentiel pendant l’anaphase.
a. Malgré la diminution progressive de leur phosphorylation, les condensines demeurent suffisamment associées aux chromatides pour préserver une structure compacte, cohérente et individualisée.
Les chromatides ne deviennent donc pas des filaments diffus, mais demeurent des entités mécaniquement organisées, capables de transmettre efficacement les forces exercées par le fuseau.
Cette organisation spatiale canalise la traction le long de l’axe des chromatides et limite les contacts désordonnés entre chromatides voisines.
b. Après la réplication en phase S, bien que les chromatides sœurs soient morphologiquement individualisées, il peut persister des entrelacements locaux entre segments d’ADN des deux chromatides, comparables à des croisements de cordes ou à de petits nœuds, appelés caténanes (Construction of a Structurally Defined Double-Stranded DNA Catenane 2011).
Lorsque ces caténanes persistent en anaphase, la structuration imposée par les condensines et la traction exercée par le fuseau soumettent ces régions à une tension mécanique croissante.
Les contraintes topologiques ne sont donc pas dissimulées dans une chromatine relâchée, mais au contraire révélées et mises en évidence par l’organisation condensine-dépendante (Origami Catenanes Templated by Gold Nanoparticles 2020)
c. Ces zones sous tension constituent des régions particulièrement accessibles à l’action de la topoisomérase IIα, qui peut y intervenir efficacement pour résoudre les derniers accrochages topologiques.
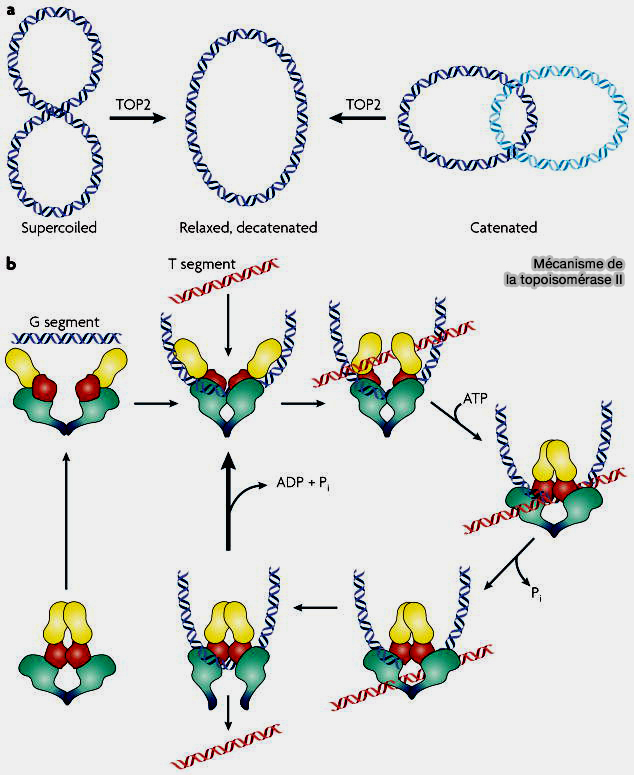
2. La topoisomérase IIα (EC 5.6.2.2), qui fonctionne en dimère, est une enzyme essentielle à la séparation physique des chromatides sœurs.
Contrairement aux condensines, qui organisent mécaniquement la chromatine, la topoisomérase IIα agit directement sur la topologie de l’ADN en modifiant sa structure par un mécanisme enzymatique actif.
Elle se fixe sur un segment d’ADN, correspondant à la première corde.
Elle crée une ouverture temporaire dans cette molécule : chaque sous-unité coupe un brin d’ADN et les deux sous-unités agissent de façon coordonnée, ce qui permet d’ouvrir transitoirement la double hélice tout en maintenant fermement les extrémités, afin de pouvoir la refermer dans l’étape suivante.
Elle permet ensuite le passage d’un autre segment d’ADN, correspondant à la deuxième corde, à travers cette ouverture, puis referme immédiatement la première molécule.
Mécanisme de la topoisomérase II
(Figure : vetopsy.fr d'après Moon et Ryu)
b. L’ADN n’est donc jamais laissé cassé dans la cellule, car la coupure est strictement transitoire et contrôlée.
Ce mécanisme permet de supprimer les croisements physiques entre molécules d’ADN et d’assurer leur séparation sans endommager leur intégrité.
4. L'activité de la topoisomérase IIα nécessite de l’énergie fournie par l’ATP, qui n’est pas utilisée pour couper l’ADN, mais pour orchestrer les mouvements de l’enzyme.
Lorsqu’elle se fixe à l’ADN, la topoisomérase adopte une conformation ouverte, qui lui permet de capturer un segment d’ADN.
La fixation de l’ATP provoque un changement de conformation de l’enzyme : elle se referme partiellement, ce qui permet de stabiliser la coupure transitoire de l’ADN et de contrôler le passage de l’autre segment à travers l’ouverture.
Après le passage du segment d’ADN, l’hydrolyse de l'ATP permet un nouveau changement de conformation qui conduit à la religature de l’ADN, puis à la libération de la molécule.
Un défaut combiné d’activité des condensines et de la topoisomérase II conduit classiquement à la persistance de ponts chromosomiques en anaphase, révélant l’échec de la résolution topologique.
Télophase
En télophase, la chute durable de l’activité des kinases mitotiques et l’action des phosphatases PP1 et PP2A entraînent une déphosphorylation généralisée des sous-unités des condensines, ce qui réduit progressivement leur affinité pour la chromatine et favorise leur dissociation.
1. La perte des complexes condensines actifs s’accompagne d’un désassemblage progressif de l’architecture chromosomique mitotique.
La diminution de la densité de bouclage conduit à une relaxation graduelle de la compaction, la disparition de l’organisation hélicoïdale et la perte de la rigidité mécanique caractéristique des chromatides.
2. La reformation de l’enveloppe nucléaire et la reprise des fonctions nucléaires interphasiques (transcription, réplication, organisation spatiale du génome) s’accompagnent d’une transition vers une organisation chromatinienne dominée par les interactions locales, les TAD (Topologically Associating Domain) et les compartiments A/B, compatibles avec l’expression génique et la dynamique nucléaire.
Les condensines retrouvent alors leur état de distribution interphasique, la condensine II redevient principalement nucléaire mais faiblement associée à la chromatine, la condensine I redevient majoritairement cytoplasmique.
Remarque : les défauts d’expression, d’assemblage ou de régulation des condensines sont associés à une instabilité chromosomique, à des défauts de condensation et de ségrégation, ainsi qu’à une augmentation des erreurs mitotiques.
Ces altérations sont impliquées dans divers contextes pathologiques, notamment certaines aneuploïdies constitutionnelles, de nombreux cancers et plusieurs syndromes développementaux rares liés à des mutations des sous-unités des complexes condensines.
Ces observations cliniques et génétiques soulignent le rôle central des condensines dans le maintien de l’intégrité du génome et la stabilité des divisions cellulaires.