La métaphase, qui dans la mitose suit la prométaphase, correspond à l’alignement complet des chromosomes sur la plaque équatoriale sous l’effet des forces bipolaires du fuseau.
La métaphase correspond à l’établissement d’un état d’équilibre mécanique du fuseau mitotique, caractérisé par l’alignement complet des chromosomes sur la plaque équatoriale.
La métaphase est caractérisée par l’alignement de l’ensemble des chromosomes sur la plaque équatoriale, une position d’équilibre dynamique définie par l’application de forces de traction symétriques sur les centromères par les microtubules kinétochoriens issus des deux pôles.
La plaque équatoriale ne correspond pas à une structure matérielle, mais à une position dynamique d’équilibre et constitue ainsi le point de référence spatial de la métaphase et le marqueur de la stabilisation mécanique préalable à l’entrée en anaphase.
Attachements end-on stables
1. L’alignement métaphasique repose sur l’établissement d’attachements end-on stables entre les kinétochores et les microtubules du fuseau.
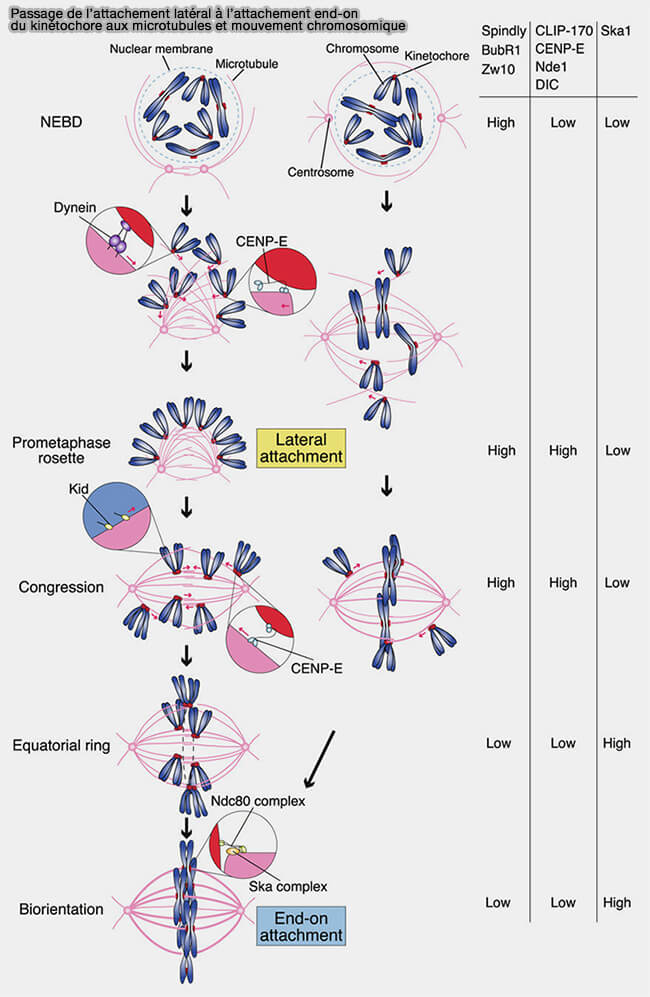
Il s’oppose à l’attachement latéral, de nature transitoire, dans lequel le kinétochore interagit avec la paroi du microtubule via des moteurs moléculaires recrutés par la corona fibreuse, notamment la kinésine CENP-E et la dynéine, permettant un glissement exploratoire sans couplage à l’extrémité (+) ni génération de tension bipolaire stable.
b. L’attachement end-on permet :
la transmission directe des forces de polymérisation/dépolymérisation,
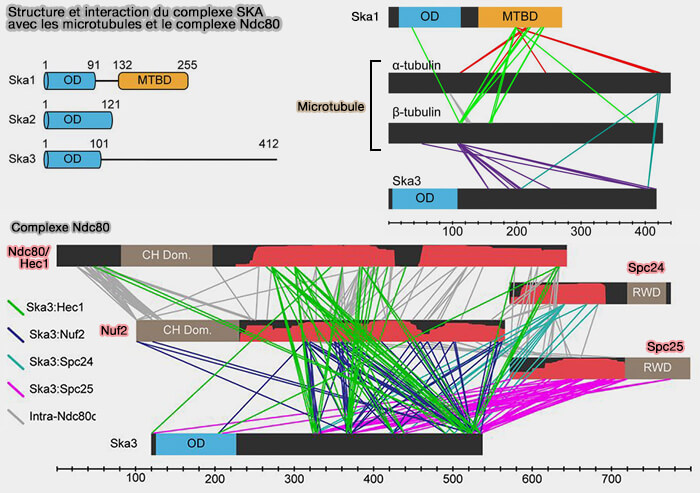
Ska1, Ska2 et Ska3 forment ensemble une structure stable capable de s’associer à la fois au réseau KMN et aux microtubules.
Le recrutement du complexe SKA au kinétochore est dépendant du complexe Ndc80 qui permet à SKA de s’associer aux microtubules via des interfaces qui favorisent son accumulation le long des protofilaments.
Structure et interactions de SKA
(Figure : vetopsy.fr d'après Helgeson et coll)
SKA3 possède une queue C-terminale disordonnée qui interagit avec les régions coiled-coil de chaque sous-unité du complexe NDC80, créant une liaison coopérative qui renforce l’interface kinétochore-microtubule.
SKA2 est une sous-unité d’assemblage du complexe qui contribue à la stabilité de l’hétéro-oligomère et à son organisation correcte.
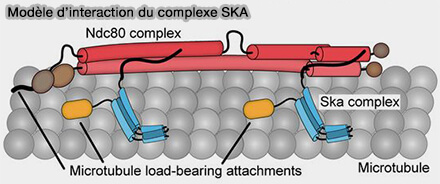
b. Le complexe SKA renforce la connexion entre le kinétochore et les extrémités des microtubules, en particulier lors de phases de dépolymérisation.
Modèle d'interaction de SKA
(Figure : vetopsy.fr d'après Helgeson et coll)
Il supporte des forces appliquées à l’extrémité des microtubules, ce qui renforce l’attachement de Ndc80 dans des conditions de tension mécanique.
Il favorise le maintien du contact avec les microtubules en dépolymérisation, transformant ainsi les variations de longueur des microtubules en forces de traction que le kinétochore peut utiliser pour maintenir la tension.
En coopérant avec Ndc80, SKA augmente la durée de vie des attachements end-on, réduisant les détachements intempestifs et contribuant à la précision de la bi-orientation.
Biorientation et positionnement équatorial des chromosomes
1. La biorientation correcte des kinétochores assure la stabilité mécanique des attachements ( fuseau en métaphase).
Chaque paire de chromatides sœurs établit des attachements end-on stables avec des microtubules issus de pôles opposés, via le réseau KMN, ce qui limite les glissements latéraux et renforce la précision du positionnement.
Cette configuration sous tension constitue le critère mécanique central distinguant les attachements corrects des attachements erronés encore susceptibles d’être dissociés, et conditionne la levée ultérieure du SAC.
2. La position équatoriale des chromosomes résulte d’un équilibre dynamique des forces au sein du fuseau par la combinaison de plusieurs forces.
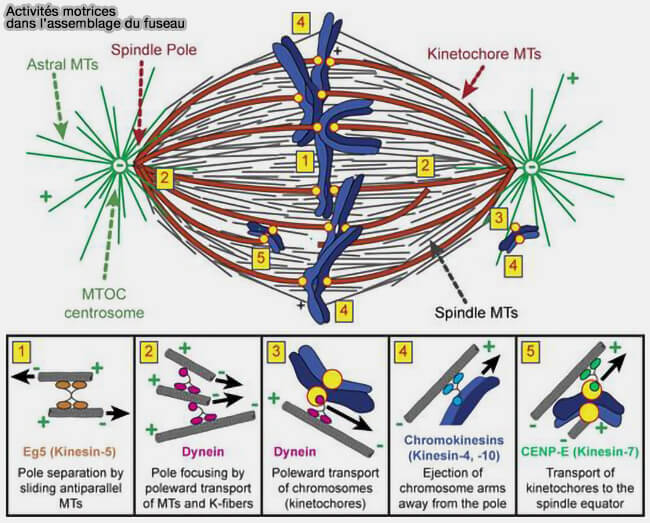
c. Les forces de poussée exercées sur les bras chromosomiques par certaines kinésines chromosomiques contribuent également à ce processus.
KIF22 (Kid, kinésine-10) génère une force polaire d’éjection repoussant les chromosomes loin des pôles.
Cette force s’oppose à la traction kinétochorienne et contribue au centrage des chromosomes sur la plaque équatoriale, sans intervenir directement dans les attachements end-on des kinétochores.
KIF4A (kinésine-4) n’induit pas une poussée supplémentaire mais module l’intensité et la distribution spatiale, de cette force d’éjection, en limitant les fluctuations excessives et en affinant la stabilité du positionnement métaphasique.
KIF4A intervient aussi comme régulateur de l’architecture de la zone médiane en limitant l’allongement excessif des microtubules dans la zone de chevauchement.
Activités motrices dans l'assemblage du fuseau
(Figure : vetopsy.fr d'après Heald et Khodjakov)
3. Une fois la biorientation correcte établie et la tension bipolaire stabilisée en métaphase, le fuseau mitotique quitte progressivement un état d’équilibre dédié au positionnement des chromosomes, préparant la transition vers les dynamiques d’écartement des pôles caractéristiques de l’anaphase.
Remarque : la dynamique des microtubules kinétochoriens est finement régulée par l’instabilité dynamique de leurs extrémités (+), permettant des cycles coordonnés de polymérisation et de dépolymérisation qui ajustent en permanence les forces exercées sur chaque chromosome et corrigent les fluctuations avant l’engagement de la séparation des chromatides.
2. La tension exercée sur les kinétochores module l’activité des kinases et stabilise les attachements.
a. La mise sous tension des chromatides sœurs réduit l’activité corrective d’Aurora B, positionnée au sein du centromère et sensible à l’absence de tension.
Lorsque la tension est suffisante, les substrats kinétochoriens s’éloignent du gradient d’Aurora B, ce qui limite la phosphorylation déstabilisante des composants du complexe KMN et stabilise les attachements microtubule-kinétochore.
b. Cette stabilisation mécanique constitue le signal principal permettant de distinguer les attachements corrects des attachements erronés encore susceptibles d’être dissociés.
Elle s’accompagne du recrutement préférentiel de complexes associés aux attachements stables, notamment le complexe Astrin-SKAP, qui s’accumule sur les kinétochores correctement attachés et sous tension et contribue à verrouiller les attachements end-on.
Aurora A agit principalement au niveau des pôles pour contrôler la dynamique des microtubules polaires et maintenir la symétrie du fuseau bipolaire.
Plk1 renforce la maturation fonctionnelle des pôles, stabilise les interactions kinétochore-microtubule et contribue au maintien de l’architecture bipolaire indispensable à l’alignement équatorial.
Tant qu’un kinétochore demeure imparfaitement stabilisé ou insuffisamment tendu, le SAC maintient une production résiduelle de complexe MCC (Mitotic Checkpoint Complex), empêchant l’activation complète de l’APC/CCdc20.
Cette surveillance moléculaire garantit que toute anomalie tardive d’attachement ou de tension soit immédiatement détectée, évitant une entrée prématurée en anaphase.
Lorsque l’ensemble des kinétochores présente des attachements biorientés stables et une tension homogène, la production de MCC cesse et Cdc20 est progressivement libéré.
Passage à l'anaphase
La transition vers l’anaphase débute lorsque l’APC/CCdc20 devient pleinement actif, déclenchant la dégradation de la sécurine et permettant l’activation de la séparase, ce qui conduit à la séparation synchronisée des chromatides sœurs.




 forces dans le fuseau mitotique),
forces dans le fuseau mitotique),