


Cycle cellulaire
Reproduction cellulaire
Kinétochore et attachements chromosomiques
Corona fibreuse
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

La corona fibreuse est une structure périphérique transitoire du kinétochore non attaché, spécialisée dans la capture initiale des microtubules du fuseau et l’amplification du point de contrôle du fuseau (SAC).
Corona fibreuse : plateforme de capture et signalisation
La corona fibreuse (fibrous corona) est une couche périphérique et transitoire située à la périphérie de la couche externe, au‑delà du réseau KMN.

La corona fibreuse est présente uniquement sur les kinétochores non attachés aux microtubules, principalement en prométaphase, et se résorbe dès que les attachements end-on stables et la tension bipolaire sont établis.
Composition de la corona fibreuse
1. La corona fibreuse (fibrous corona) est composée par :
- le complexe RZZ (ROD/KNTC1-ZW10-ZWILCH),
- Spindly, adaptateur du RZZ,
- le moteur dynéine-dynactine,
- le complexe Mad1-Mad2,
- d’autres composants du point de contrôle du fuseau (SAC), notamment BubR1:Bub3, recrutés à la corona en coordination avec la plateforme KNL1-Bub1 (Distinct domains in Bub1 localize RZZ and BubR1 to kinetochores to regulate the checkpoint 2015).
2. Cette organisation forme une matrice périphérique étendue, capable d’augmenter la surface fonctionnelle du kinétochore non attaché.
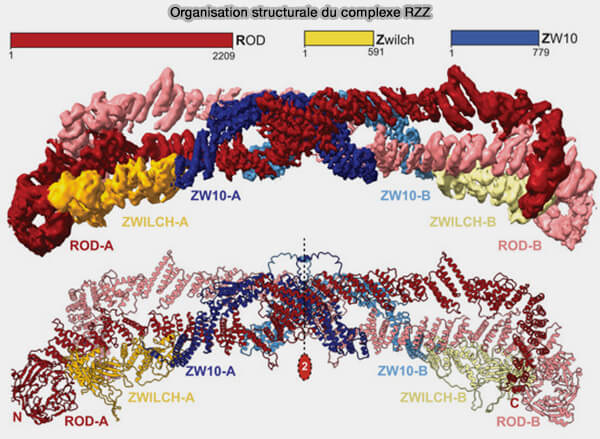
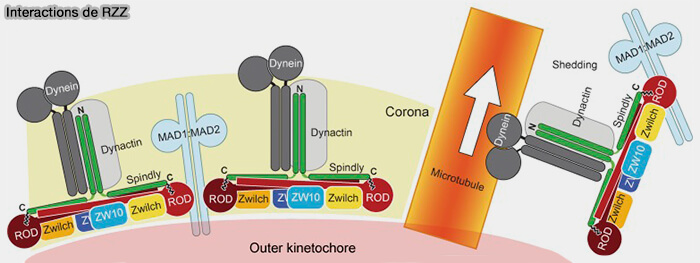
Structure du complexe RZZ
Le complexe RZZ, noyau structural et fonctionnel de la corona, est un assemblage tripartite rigide mais dynamique, qui coordonne le recrutement des principaux effecteurs du point de contrôle du fuseau (SAC), ainsi que des moteurs impliqués dans sa désactivation (Structure of the RZZ complex and molecular basis of Spindly‐driven corona assembly at human kinetochores 2022).
Le complexe RZZ forme des dimères stables, qui constituent l’unité minimale de son assemblage, et possède une capacité intrinsèque d’oligomérisation permettant la formation de la matrice étendue de la corona fibreuse.
(Figure : vetopsy.fr d'après Raisch et coll)
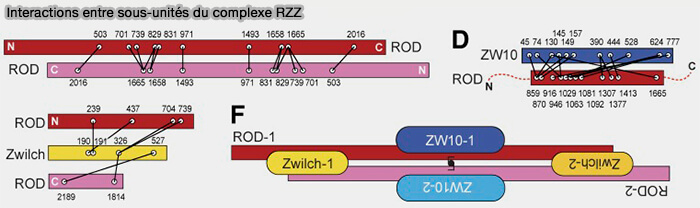
1. ROD/KNTC1 (Rough Deal), élément central du complexe, est une protéine de grande taille, majoritairement alpha-hélicoïdale, qui forme une charpente allongée servant de support à l’assemblage de ZW10 et de ZWILCH qui assure :
- l’architecture globale du complexe RZZ,
- la capacité d’auto-assemblage et d’expansion de la corona fibreuse,
- le rôle de plateforme structurale pour le recrutement des partenaires du point de contrôle du fuseau (SAC).
2. ZW10 s’associe étroitement à ROD et contribue à la stabilisation du complexe RZZ en participant :
- à l’organisation de l’interface du complexe avec le module dynéine-dynactine,
- au couplage fonctionnel du RZZ au module Spindly-dynéine-dynactine, sans constituer à elle seule un adaptateur moteur.
(Figure : vetopsy.fr d'après Mosalagenti et coll)
3. ZWILCH agit comme sous-unité régulatrice en :
- contribuant à l’organisation et à la reconnaissance des partenaires du SAC au sein de la corona,
- participant à la stabilité et à la fonctionnalité du complexe RZZ dans un contexte dépendant de l’état d’attachement du kinétochore.
Recrutement des autres modules de la corona fibreuse
Le complexe RZZ constitue la plateforme centrale de recrutement des composants périphériques de la corona fibreuse en organisant l’assemblage de modules adaptateurs et moteurs nécessaires aux fonctions ultérieures de la corona, sans les activer directement à ce stade.
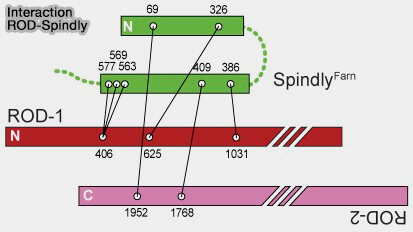
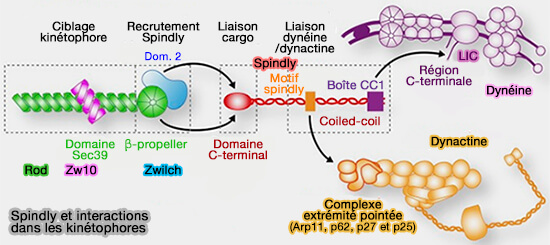
1. Dans la corona fibreuse, le complexe RZZ recrute Spindly, pour former un module RZZ-Spindly-dynéine, dans lequel Spindly joue le rôle d’adaptateur moléculaire (RZZ‐Spindly and CENP‐E form an integrated platform to recruit dynein to the kinetochore corona 2023).
(Figure : vetopsy.fr d'après Mosalagenti et coll)
a. Après la farnésylation dépendante de son motif C-terminal CAAX, Spindly se lie directement à ROD par des interactions multiples, ce qui permet son ancrage stable à la corona fibreuse et la formation d’une plateforme structurale propice à l’arrimage de la dynéine-dynactine.
b. Spindly recrute directement le complexe dynéine-dynactine via des motifs conservés, permettant l’engagement fonctionnel du moteur au niveau de la corona fibreuse (Molecular mechanism of dynein recruitment to kinetochores by the Rod-Zw10-Zwilch complex and Spindly 2017).
- Sur le plan moléculaire, Spindly agit comme un adaptateur de la dynéine-dynactine en recrutant directement le complexe moteur via un motif Spindly conservé, qui interagit avec les sous-unités p25/p27 de la dynactine, tandis que ses régions coiled-coil contribuent à l’activation et à la processivité du moteur au kinétochore (
 Spindly et dynactine).
Spindly et dynactine). - Sur le plan mécanistique, Spindly est décrite comme un adaptateur de type BicD2-like, favorisant l’assemblage productif du complexe dynéine-dynactine-adaptateur et l’activation du transport rétrograde des composants du SAC (Removal of Spindly from microtubule-attached kinetochores controls spindle checkpoint silencing in human cells 2010).
(Figure : vetopsy.fr d'après Gama et coll)
c. À ce stade, la dynéine est recrutée et positionnée, mais son activité motrice reste conditionnée par l’état d’attachement et de tension du kinétochore (Structure of the RZZ complex and molecular basis of its interaction with Spindly 2017).
2. Le complexe RZZ participe également au recrutement du complexe Mad1-Mad2 à la corona fibreuse ( activation et amplification du SAC).
(Figure : vetopsy.fr d'après Mosalagenti et coll)
Rôle de la corana fibreuse
Le recrutement du complexe RZZ positionne les composants du SAC sur les kinétochores non attachés et prépare leur activation fonctionnelle.
Le point de contrôle du fuseau (SAC) est étudié dans des chapitres spécifiques.
Activation et amplification du SAC
L’activation du point de contrôle du fuseau (SAC) repose sur la capacité des kinétochores non attachés à générer et amplifier un signal inhibiteur empêchant l’activation prématurée de l’APC/C.
1. En prométaphase, la présence d’une corona fibreuse pleinement développée permet le recrutement et le maintien du complexe Mad1-Mad2 à la périphérie du kinétochore.
- Il n’existe pas de liaison directe Mad1-RZZ bien définie comme CENP-C/Mis12 ou Ndc80/microtubule.
- L’association de Mad1-Mad2 à la corona fibreuse est médiée par un ensemble d’interactions faibles, multivalentes et coopératives, organisées par le complexe RZZ.
a. ROD (KNTC1) constitue la charpente principale de la corona fibreuse et sa déplétion entraîne une perte majeure de Mad1-Mad2 aux kinétochores, car Mad1 ne peut pas être maintenue à la périphérie du kinétochore.
b. ZWILCH agit comme sous-unité régulatrice, contribuant à la rétention locale de Mad1 au sein de la matrice RZZ.
c. Spindly n’est pas un ligand direct majeur de Mad1, mais agit selon deux modalités complémentaires :
- d'une part, par sa liaison à ROD/ZWILCH qui stabilise l’architecture de la corona, ce qui favorise le maintien de Mad1-Mad2 dans cette structure.
- d'autre part, par sa liaison au module dynéine-dynactine qui permet une rétention stable mais réversible de Mad1 au sein de la corona.
Lorsque l’attachement microtubulaire et la tension bipolaire sont établis, cette même architecture facilite le basculement rapide vers le transport rétrograde des composants du SAC, expliquant l’efficacité du retrait de Mad1 lors de la levée du checkpoint.
(Figure : vetopsy.fr d'après Allan et coll)
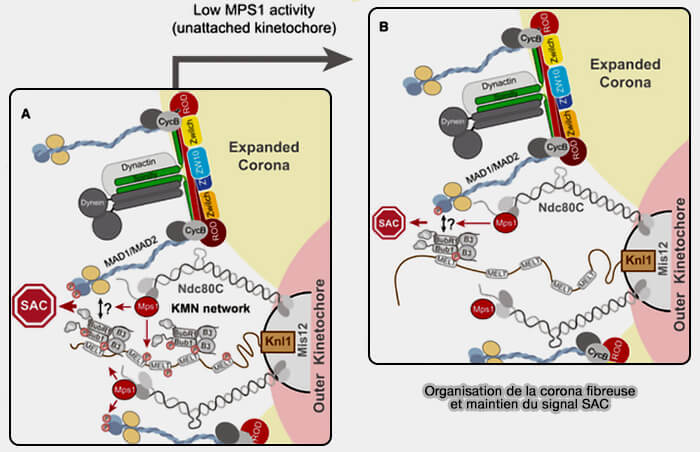
2. En complément de cette organisation coronale, la cycline B1, principalement sous forme de complexe cycline B/CDK1 (MPF), contribue au maintien fonctionnel du signal SAC au niveau du kinétochore (figure A de Cyclin B1 scaffolds MAD1 at the kinetochore corona to activate the mitotic checkpoint 2020).
a. Elle s’accumule à la corona fibreuse où elle interagit avec Mad1, renforçant sa rétention locale et permettant la persistance d’un signal SAC robuste, même lorsque l’activité de MPS1 diminue (Figure B).
- Cette fonction d’échafaudage ne repose pas sur une activation primaire du checkpoint, mais sur le maintien d’un environnement mitotique permissif au signal SAC.
- Par ce mécanisme, la cycline B1 établit un lien direct entre l’état du checkpoint et la régulation de l’APC/C (Anaphase-Promoting Complex/Cyclosome).
b. Lors de l’établissement de l’attachement microtubulaire correct et de la tension bipolaire, la disparition progressive de la cycline B1 du kinétochore accompagne la résorption de la corona fibreuse et contribue à la levée du SAC.
2. Le recrutement de Mad1-Mad2 à la corona fibreuse par le complexe RZZ est une voie largement indépendante de la plateforme KNL1-Bub1, conférant au SAC une redondance fonctionnelle.
Cette organisation sécurise l’activation du checkpoint en début de mitose et permet le maintien du signal tant que l’attachement correct aux microtubules n’est pas établi, évitant une levée prématurée du contrôle en cas d’attachements instables ou transitoires.
a. Une fois positionné à la corona, Mad1 stabilise Mad2 et catalyse sa conversion de la conformation ouverte (O-Mad2) vers la conformation fermée (C-Mad2) pour s’incorporer au complexe MCC (Mitotic Checkpoint Complex) en association avec BubR1, Bub3 et Cdc20, qui inhibe l’APC/CCdc20 et l’entrée prématurée en anaphase.
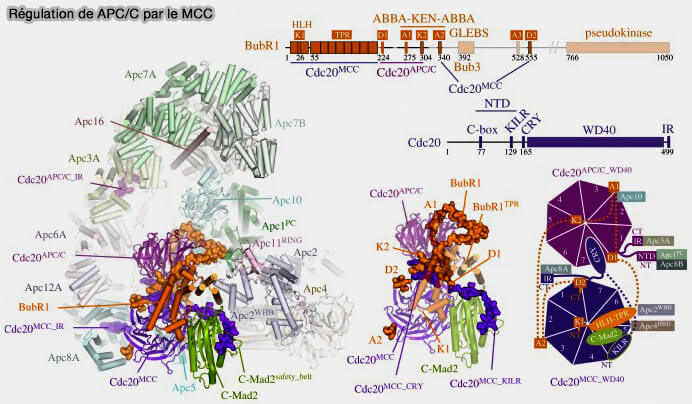
(Figure : vetopsy.fr d'après Alfieri et coll)
b. La localisation périphérique du complexe Mad1-Mad2 au sein de la corona fibreuse augmente la surface et la durée de signalisation sur les kinétochores non attachés.
Rôle dans la capture initiale des microtubules
En prométaphase, l’expansion de la corona fibreuse augmente considérablement la surface d’interaction du kinétochore avec le fuseau mitotique.
Cette structure périphérique étendue favorise la capture initiale des microtubules par des interactions latérales, avant l’établissement d’un attachement end-on stable médié par le complexe NDC80.
1. L’attachement latéral correspond à une interaction transitoire entre la paroi du microtubule et la surface du kinétochore, principalement médiée par des moteurs moléculaires et des complexes périphériques de la corona fibreuse.
On emploie l'expression attachement latéral car le kinétochore interagit avec la paroi du microtubule, et non avec son extrémité (+) comme dans l'attachement end-on, ce qui confère à cette interaction une géométrie glissante, réversible et dépourvue de tension bipolaire stable (Finding the middle ground: how kinetochores power chromosome congression 2010).
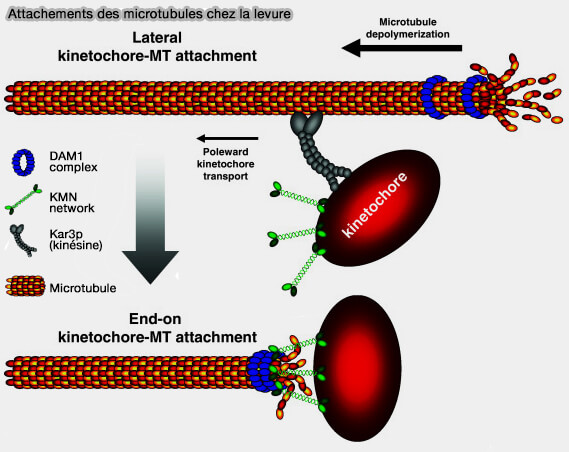
(Figure : vetopsy.fr d'après Kops et coll)
Remarque : sur la figure ancienne ci-dessus, les protéines Dam1 et Kar3p correspondent à des composants spécifiques des levures, dont les fonctions, chez les métazoaires,, sont assurées par le complexe Ndc80, le complexe SKA et des moteurs tels que la dynéine et CENP-E.
Vous pouvez voir les différences entre levure et métazoaires dans : Kinetochore Architecture Employs Diverse Linker Strategies Across Evolution (2022).
2. La corona fibreuse agit comme une zone de prospection dynamique, permettant aux kinétochores encore éloignés du plan équatorial ou mal orientés d’entrer en contact avec les microtubules du fuseau.
a. Sur le plan moléculaire, cet attachement repose notamment sur :
- la dynéine-dynactine, recrutée via le module RZZ-Spindly, qui permet la capture initiale des microtubules et le déplacement du chromosome vers le pôle,
- la kinésine CENP-E, qui assure le glissement directionnel du kinétochore le long de la paroi du microtubule et favorise la conversion ultérieure vers un attachement end-on,
- des composants de la corona fibreuse (RZZ, Mad1-Mad2, BubR1:Bub3), qui augmentent la surface de contact et stabilisent temporairement ces interactions latérales.
b. Mécaniquement, l’attachement latéral permet :
- la capture exploratoire des microtubules du fuseau,
- le glissement du kinétochore le long du microtubule sans couplage direct à l’extrémité (+),
- une interaction faiblement résistante à la force, réversible et dépourvue de tension bipolaire stable.
c. Ce mécanisme est particulièrement important pour les chromosomes initialement mal positionnés, pour lesquels une capture directe end-on serait peu probable.
3. Une fois l’attachement end-on établi via le complexe Ndc80 et la tension bipolaire générée, le rôle de la corona fibreuse dans la capture devient obsolète, et sa résorption accompagne la transition vers un kinétochore pleinement engagé dans la ségrégation chromosomique.
Remarque : les complexes RZZ et Astrin-SKAP définissent deux états mutuellement exclusifs du kinétochore :
- un état exploratoire et signalétique (RZZ, SAC actif),
- un état stabilisé et permissif (Astrin-SKAP, SAC levé), assurant la transition ordonnée vers la métaphase stable et l’entrée en anaphase.
Désassemblage de la corona fibreuse et levée du SAC
La levée du point de contrôle du fuseau (SAC) est un processus actif, étroitement couplé à l’établissement correct des attachements microtubulaires et à la génération de tension bipolaire.
1. À la transition prométaphase/métaphase (transition G2/M), lorsque les microtubules établissent un attachement end-on stable avec le complexe NDC80 et que la tension bipolaire est appliquée, l’architecture du kinétochore se réorganise.
a. Ces changements limitent l’accès des kinases au checkpoint aux plateformes de signalisation et initient la rétraction de la corona fibreuse.
- MPS1 dépend de l’accessibilité du domaine CH de Ndc80, de l’attachement end-on et de la tension qui empêche sa fixation, provoque l'arrêt de la phosphorylation des motif MELT de KNL1.
- Aurora B, au sein du CPC (Chromosomal Passenger Complex), reste centromérique et l’augmentation de la distance centromère-kinétochore sous tension réduit son action sur les substrats kinétochoriens et initie la rétraction de la corona fibreuse.
b. À ce stade, les composants du SAC, à, savoir Mad1-Mad2, BubR1, Bub3, ainsi que le complexe RZZ, Spindly et la dynéine, restent encore transitoirement associés à la corona, mais leur maintien devient instable.
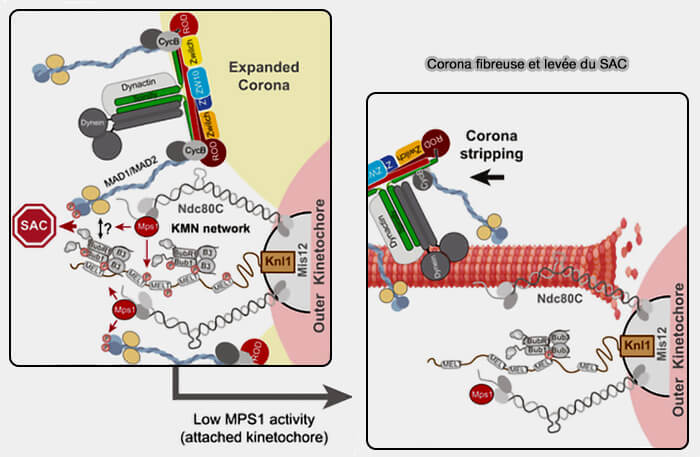
(Figure : vetopsy.fr d'après Allan et coll)
2. La dynéine, préalablement recrutée à la corona fibreuse via le module RZZ-Spindly en prométaphase, devient alors fonctionnellement active et engage un transport rétrograde le long des microtubules du fuseau, dirigeant les composants du SAC vers les pôles (Cyclin B1 scaffolds MAD1 at the kinetochore corona to activate the mitotic checkpoint 2020).
a. Ce mécanisme, souvent désigné sous le terme de stripping, en littéralement dépouillement ou retrait, correspond à l’élimination physique de (Dynein at kinetochores: Making the connection 2017) :
- Mad1-Mad2, BubR1 et Bub3,
- Spindly,
- une fraction des composants du complexe RZZ, étroitement associés à Spindly, sont partiellement entraînés dans ce transport, tout en se désassemblant progressivement de la couche externe.
b. Ce transport motorisé entraîne la désorganisation progressive de la corona fibreuse et constitue une étape déterminante de l’extinction du signal checkpoint au niveau des kinétochores correctement attachés.
c. L'activation de la dynéine est conditionnée par l’état mécanique du kinétochore.
- L’établissement d’un attachement end-on stable entre les microtubules et le complexe Ndc80, associé à la génération de tension bipolaire, entraîne une réorganisation de l’architecture kinétochorienne et le début de la rétraction de la corona fibreuse.
- Ces changements s’accompagnent d’une réduction locale de l’activité des kinases du checkpoint, notamment MPS1, et d’une déphosphorylation progressive des plateformes SAC, diminuant l’ancrage des complexes Mad et Bub à la corona.
- Parallèlement, les modifications conformationnelles induites par la tension favorisent une interaction productive entre Spindly et le complexe dynéine-dynactine, permettant l’activation effective du moteur.
d. Cette activation de la dynéine, combinée à la perte des signaux de maintien du checkpoint et aux contraintes mécaniques exercées sur le kinétochore, entraîne la désorganisation progressive du complexe RZZ, dont une fraction est cotransportée lors du stripping tandis que le reste se dissocie localement, conduisant à la résorption complète de la corona fibreuse.
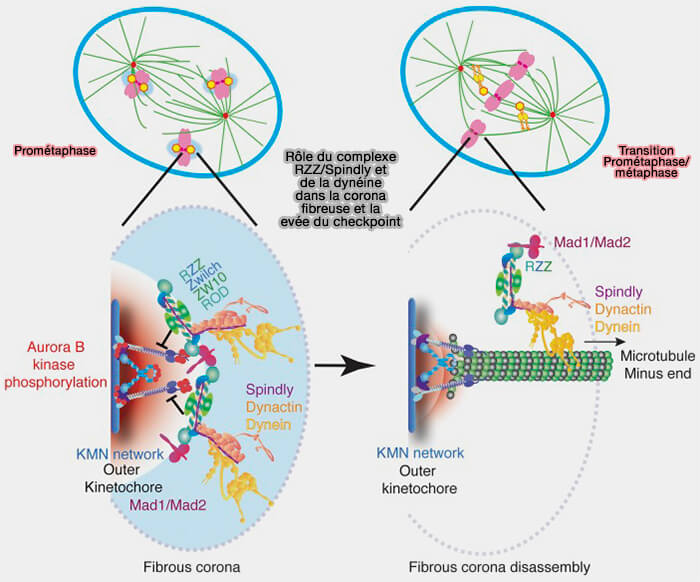
(Figure : vetopsy.fr d'après McHugh et Welburn)
3. La disparition de cette structure périphérique marque le retour à une organisation du kinétochore dominée par le cœur du réseau KMN, désormais engagé dans des attachements microtubulaires stables et sous tension.
4. La levée complète du SAC survient une fois l’ensemble des kinétochores correctement attachés et libérés de leurs signaux checkpoint, ce qui permet l’engagement de la transition métaphase-anaphase.
L’activation de l’APC/CCdc20 est ainsi obtenue par un double mécanisme coordonné :
- diminution de la signalisation liée à l’attachement et à la tension,
- élimination active des complexes inhibiteurs par la dynéine.
Régulation du kinétochore
Biologie cellulaire et moléculaireConstituants de la celluleReproduction cellulaireMatériel génétiqueFuseauCycle cellulaireInterphasePhase G1Phase SPhase G2MitoseProphasePrométaphaseMétaphaseAnaphaseTélophaseCytokinèseAbcissionMéioseRégulation du cycle cellulaireCyclinesCDKComplexes Cyclines/CDKPoints de contrôles (checkponts)Enzymes mitotiquesMoteurs mitotiquesComplexes SMC (condensines et cohésines)BiochimieTransport membranaire Moteurs moléculairesVoies de signalisation