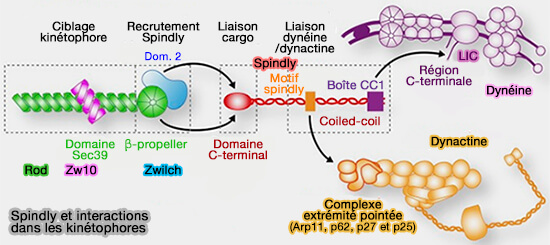
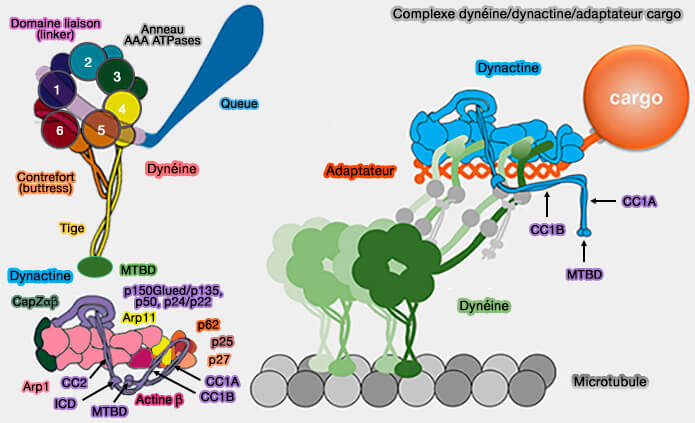
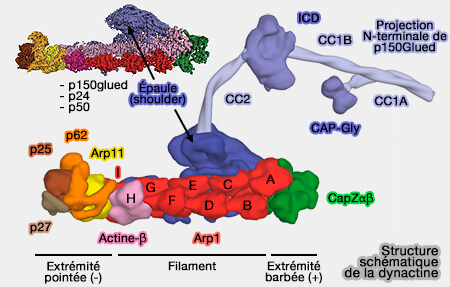
1. Le filament de dynactine, i.e. domaine Arp1, comprend :
Structure de la dynactine
(Figure : vetopsy.fr d'après plusieurs sources)
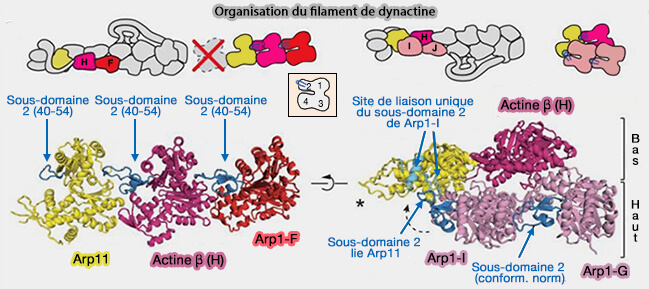
8 molécules d'Arp1 (Actin-related protein 1 ou ACRTR1A), formant deux protofilaments, i.e. le protofilament supérieur en contient 5 (A,C, E, G, I) et le protofilament inférieur 3 (B,D, F),
d'une molécule de β-actine qui se place en H, i.e. en-dessous de I et G, à côté de F.
Arp1 et la β-actine ont 53 % d'identité de séquence ( structure de Arp1).
Ils sont formés de quatre sous-domaines entourant un site de liaison de nucléotide.
Un contact clé dans le filament est la boucle du sous-domaine 2 qui peut interagir avec le sillon des sous-domaines 1 et 3 de l'unité voisine (cf. figure suivante).
2. À l'extrémité pointée (-), le protofilament inférieur se termine par la β-actine (H) tandis que le supérieur se termine par une unité Arp1, i.e. Arp1-I, en contact avec Arp11 ( Arp11 et extrémité pointée).
Remarque : certains auteurs incluent Arp11 dans le filament de dynactine.
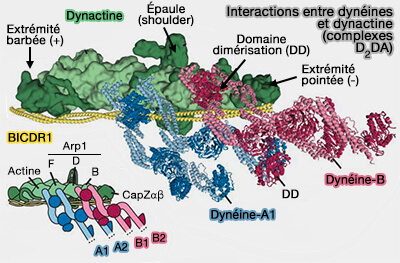
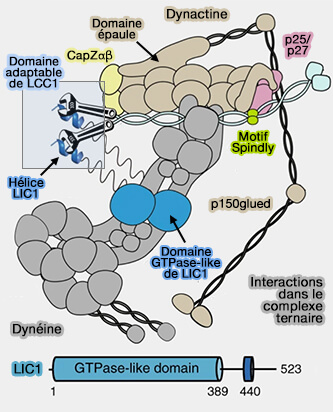
Interactions dynéine/dynactine
(Figure : vetopsy.fr d'après Urnavicius et coll)
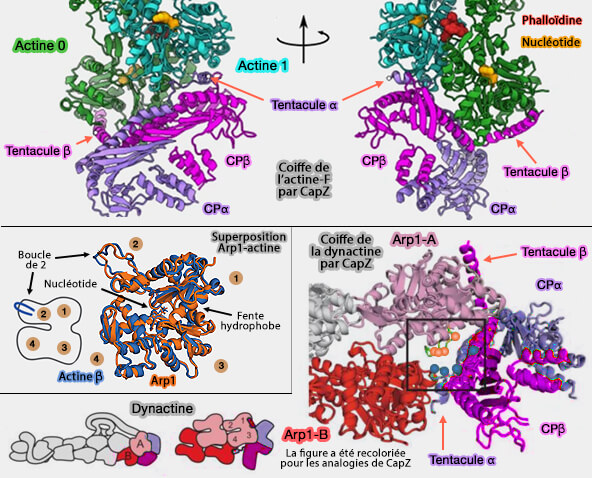
Les hélices C-terminales (tentacules) de CapZα et CapZβ s'insèrent dans les rainures des sous-domaines 1 et 3 de Arp1-A et Arp1-B, bloquant ainsi l'interaction et inhibant l'allongement des sous-unités.
La tentacule α bloque toute la surface de l'extrémité barée exposée de Arp1-B en étant insérée dans la fente hydrophobe.
La tentacule β est ancrée sur Arp1-A, mais semble conserver une certaine flexibilité comme dans l'actine.
Coiffe de l'extrémité barbée de l'actine et de la dynactine
(Figure : vetopsy.fr d'après Funk et coll et Urnavicius et coll)
Cette boucle est proche d'un groupe de quatre résidus chargés positivement sur CapZα, i.e. K223, K256, K266 et K268, suggérant que CapZαβ se lie à Arp1 avec une affinité plus élevée que l'actine.
Cela explique pourquoi CapZαβ joue un rôle direct dans la stabilisation de la structure de la dynactine.
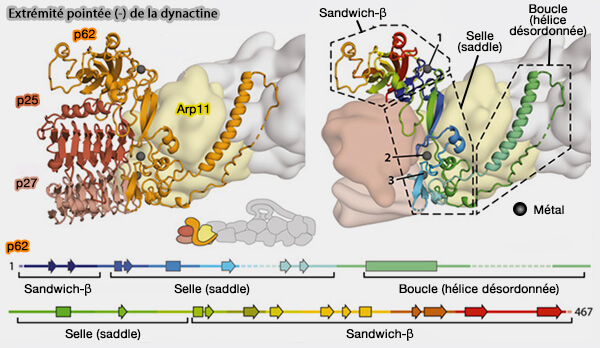
1. À l'extrémité pointée, le protofilament inférieur se termine par la β-actine (H) tandis que le supérieur se termine par une unité Arp1, i.e. Arp1-I.
Cette conformation crée un site de liaison distinctif pour Arp11.
Arp11 possède un sous-domaine-2 trop court, i.e. 14 résidus au lieu de 38, pour l'ajout de nouvelles sous-unités. au protofilament inférieur.
Arp11 se lie directement au protofilament supérieur sur la boucle du sous-domaine-2 d'Arp1-I, en bloquant stériquement l'ajout de sous-unités supplémentaires.
Arp11 peut être considéré comme la protéine de coiffe de l'extrémité pointée (-) de la dynactine.
2. Arp11 entre en contact avec p25, moins avec p27 et avec p62.
entre en contact avec la sous-unité de β-actine voisine dans le filament, i.e. par l'interaction entre K157 et K222 sur p62 à K50 et K61 sur l'actine respectivement.
1. La dynactine est un constituant des complexes DDA ou D2DA, i.e. Dynéine, Dynactine, Adaptateur.
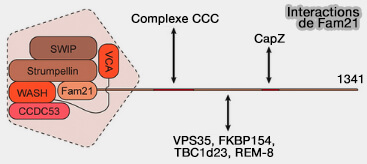
Interactions de Fam21 du complexe WASH
(Figure : vetopsy.fr d'après Wang et coll)
2. La dynactine peut être co-localisée et co-immunoprécipitée avec le complexe WASH sur la surface des membranes endosomales indiquant que les deux machines moléculaires interagissent directement ou indirectement.




 structure de Arp1).
structure de Arp1).