


Système endo-lysosomal
Complexe WASH
Complexes endosomaux
et polymérisation de l'actine
- Vue d'ensemble du système endomembranaire
- Endosomes
- Vue d'ensemble des endosomes
- Endosomes précoces
- Vue d'ensemble des endosomes précoces
- Devenirs des endosomes précoces
- Composants des endosomes précoces
- Rab5 et ses régulateurs
- Rab5 et ses effecteurs
- Complexes d'attache des endosomes précoces : complexe CORVET
- Complexe de tri et de recyclage des endosomes précoces
- Endosomes de recyclage
- Vésicules intraluminales (ILV) et endosomes ou corps mutivésiculaires (MVE/MVB)
- Endosomes tardifs
- Système endo-lysosomal et phosphoinositides
- MCS (sites de contact membranaire) RE/endosomes/lysosomes
- Lysosomes
- Trafic des endosomes/lysosomes
- Fusion et fission membranaire
- Transport membranaire
- Moteurs moléculaires
- Voies de signalisation

Le complexe WASH (Wiskott–Aldrich syndrome protein and SCAR Homologue) recrute et active le complexe Arp2/3 pour induire la polymérisation de l'actine avec l'aide d'autres complexes.
Le complexe WASH (Wiskott–Aldrich syndrome protein and SCAR Homologue) est un complexe de nucléation (NPF : Nucleation Promoting Factor) de l'actine à la surface des endosomes qui fonctionne avec d'autres complexes endosomaux.
-
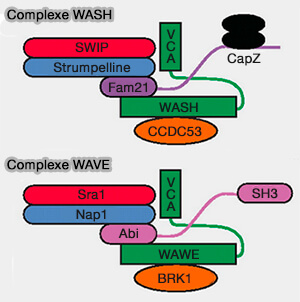
Sous-unités des complexes WASH et WAVE
(Figure : vetopsy.fr d'après Jia et coll) - Il joue un rôle clé dans la fission des tubules qui servent d'intermédiaires de transport pendant le tri des endosomes (Assembly and Activity of the WASH Molecular Machine: Distinctive Features at the Crossroads of the Actin and Microtubule Cytoskeletons 2021).
Le complexe WASH joue un rôle essentiel dans la polymérisation ramifiée de l'actine sur les endosomes, en action avec le complexe rétromère et le complexe retriever ou le complexe CCC.
Il a aussi un rôle, avec le complexe exocyste, dans l'invasion tumorale (Rôle des complexes WASH et exocyste dans l’invasion tumorale 2015).
Ces complexes sont essentiels (F-Actin Dynamics in the Regulation of Endosomal Recycling and Immune Synapse Assembly 2021) :
- pour le trafic de molécules de la membrane plasmique vers les différents compartiments récepteurs,
- pour le recyclage polarisé de molécules spécifiques vers des zones spécialisées de la cellule, comme observé, par exemple, dans les lymphocytes T subissant une synapse immunitaire (IS).
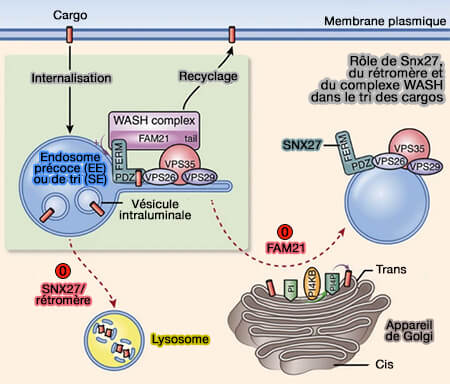
dans le tri des cargos
(Figure : vetopsy.fr d'après Lee et coll)
Complexes
endosomaux

Le complexe WASH se lie aux complexes endosomaux par VPS35.
Complexe WASH/
rétromère
1. Le complexe rétromère provoque :
- le tri et le recyclage du matériel des endosomes tardifs vers l'appareil de Golgi,
- la scission des tubules se produit aux sites de contact membranaire (MCS) avec le réticulum endoplasmique (
 MCS et fission membranaire).
MCS et fission membranaire).
2. Cette scission nécessite une protéine membranaire du RE, TMCC1 ou TransMembrane and Coiled-Coil domain family 1 qui induit le ciblage et l'accumulation aux sites de contact RE/endosome de :
- la coronine 1, la protéine liant l'actine (3),
- le complexe WASH, et, en particulier, la strumpeline (Defects in ER–endosome contacts impact lysosome function in hereditary spastic paraplegia 2018).
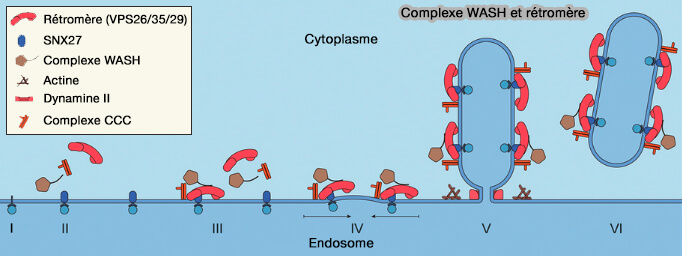
(Figure : vetopsy.fr d'après Wang et coll)
Complexe WASH/retriever
1. Le complexe retriever se compose de (Retriever, a multiprotein complex for retromer-independent endosomal cargo recycling 2017) :
-
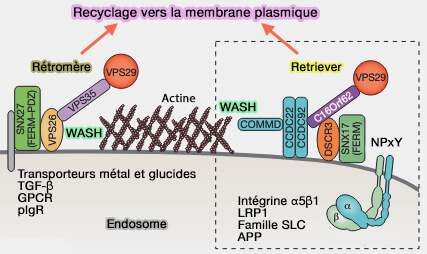
Rétromère et retriever
(Figure : vetopsy.fr d'après Rabouille) - DSCR3/Vps26C, homologue Vps26 du complexe rétromère,
- Vps29, i.e. sous-unité aussi présente dans le complexe rétromère.
2. Le complexe retriever est impliqué :
- dans le tri des cargos portant un motif NPxY/NxxY, comme les intégrines, i.e. comme le complexe retriever, dans une voie de recyclage endosomale spécialisée,
Le retriever fonctionnerait avec la nexine Snx17 pour réguler le recyclage de l'intégrine α5β1 et de plus de 120 récepteurs supplémentaires qui interagissent avec le domaine FERM de Snx17.
- dans la médiation du trafic des récepteurs des lipoprotéines de faible densité (CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL 2016).
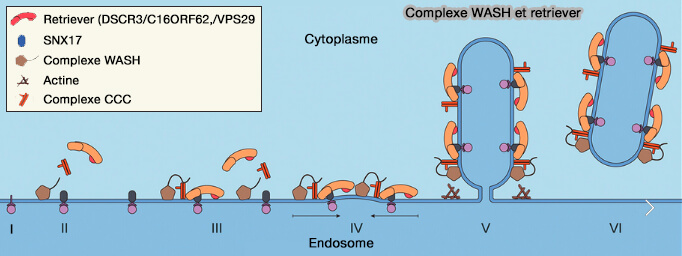
(Figure : vetopsy.fr d'après Wang et coll)
Remarque : la présence de Vps29 en tant que sous-unité à la fois dans le rétromère et le retriever, et les similitudes structurelles et fonctionnelles entre les deux complexes suggèrent une possible duplication évolutive pour élargir les capacités de régulation du recyclage à l'EE dans les organismes supérieurs ( Towards a molecular understanding of endosomal trafficking by Retromer and Retriever 2019).
Complexe WASH/CCC
1. Le complexe CCC (COMMD/CCDC22/CCDC93) comprend trois protéines :
- un membre de la famille COMMD (COMMD1-COMMD9),
- CCDC22, qui interagit avec Fam21, pour la liaison avec le complexe WASH (COMMD1 is linked to the WASH complex and regulates endosomal trafficking of the copper transporter ATP7A, 2015)
- CCDC93.
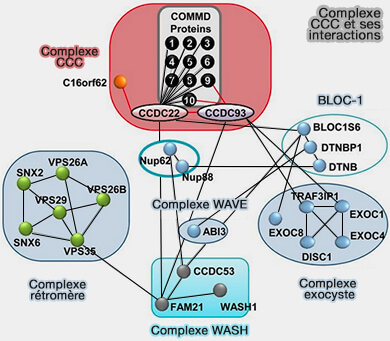
(Figure : vetopsy.fr d'après Philips-Krawczak et coll)
a. COMMD1 interagit avec CCDC93 et Vps35L, sous-unité du complexe retriever (Retriever, a multiprotein complex for retromer-independent endosomal cargo recycling 2017).
b. CCDC22 interagit avec Vps33b du complexe CHEVI (Proteomic and Biochemical Comparison of the Cellular Interaction Partners of Human VPS33A and VPS33B 2018),
Vps33B est nécessaire au recyclage des intégrines, comme dans les cellules épidermiques granulaires de la peau en interaction avec Rab11 (VPS33B and VIPAR are essential for epidermal lamellar body biogenesis and function 2018).
2. Le complexe CCC serait impliqué :
- dans le tri des cargos portant un motif NPxY/NxxY, comme les intégrines, i.e. comme le complexe retriever, dans une voie de recyclage endosomale spécialisée,
- la médiation du trafic des récepteurs des LDL (lipoprotéines de basse densité) ou LDLR (CCC- and WASH-mediated endosomal sorting of LDLR is required for normal clearance of circulating LDL 2016).
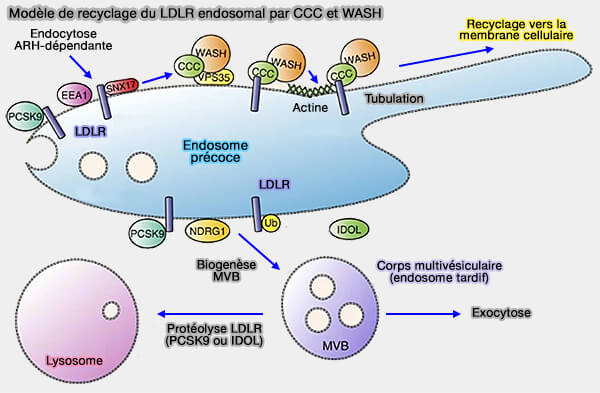
(Figure : vetopsy.fr d'après Bartuzi et coll)
Les complexes retriever et CCC avec le complexe WASH régulent aussi l'homéostasie mitochondriale (Tying trafficking to fusion and fission at the mighty mitochondria 2018).
Petites GTPases Rab et complexes endosomaux
1. Les petites GTPases, essentielles à la formation de vésicules endocytaires enveloppées, aussi bien à base de clathrine, de COPI et de COPII, sont aussi des régulateurs du trafic vésiculaire médié par le rétromère, en particulier Rab7 et TBC1D5 (petites GTPases Rab et rétromère).
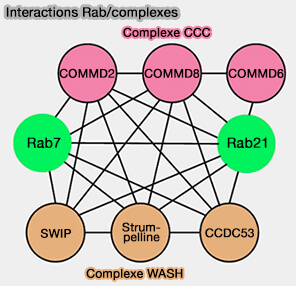
(Figure : vetopsy.fr d'après Clague et Urbé)
2. Bien que l'on sache peu de choses sur l'interaction entre les Rab GTPases et les complexes retriever et CCC et WASH, une étude protéomique récente axée sur les protéines SNARE et les petites GTPase Rab a identifié des clusters fonctionnels, comme une corrélation entre Rab10 et la syntaxine4 (Stx4) ou entre Rab7/Rab21 et les complexes WASH et CCC (Data Mining for Traffic Information 2020).
Mode d'action du complexe WASH
Vue d'ensemble
1. L'actine ramifiée sur les endosomes est dépendante du complexe WASH.
WASH contribue directement au tri des récepteurs à condition qu'ils aient une affinité pour l'actine (HRS–WASH axis governs actin-mediated endosomal recycling and cell invasion 2018).
L'actine ramifiée maintient les microdomaines lipidiques dans la membrane endosomale, surtout enrichis en Rab5.
L'altération de la polymérisation de l'actine provoque la fusion de ces microdomaines en un seul, le complexe WASH est alors incapable de le détacher de la membrane endosomale (Actin Polymerization Controls the Organization of WASH Domains at the Surface of Endosomes 2012).
2. Le complexe WASH est localisé à la base des tubules endosomaux, i.e. au bon endroit pour effectuer la scission (The Arp2/3 Activator WASH Controls the Fission of Endosomes through a Large Multiprotein Complex 2009).
Les endosomes sont des structures tubulo-vésiculaires, et la traction des membranes endosomales par les moteurs des microtubules, i.e. dynéine et kinésines, se poursuit lorsque la formation d'actine ramifiée est altérée.
3. La dynactine peut être co-localisée et co-immunoprécipitée avec le complexe WASH, indiquant que les deux machines moléculaires interagissent directement ou indirectement (The Arp1/11 minifilament of dynactin primes the endosomal Arp2/3 complex 2021).
La dynactine, adaptateur essentiel pour le transport médié par la dynéine, est étudiée dans un chapitre spécial.
Mécanisme probable
Le complexe WASH et la dynactine sont à proximité.
- Le complexe WASH est recruté à la surface endosomale par l'interaction de FAM21 avec la sous-unité du rétromère VPS35 du rétromère, mais aussi avec SNX27.
- La dynactine interagit avec les nexines de tri SNX-BAR, i.e. SNX1/2 et SNX5/6.
Le mécanisme semble être le suivant pour le rétromère (The Arp1/11 minifilament of dynactin primes the endosomal Arp2/3 complex 2021).
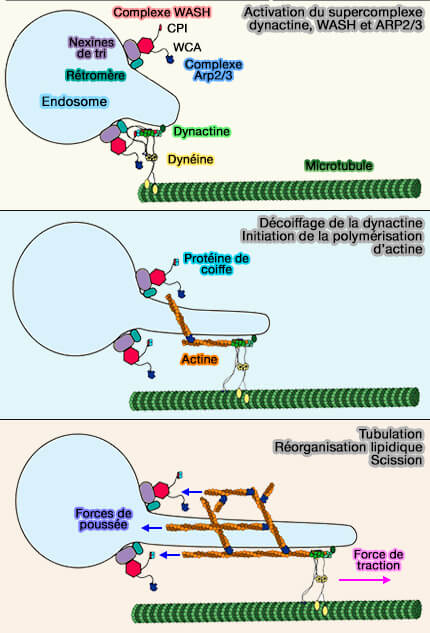
(Figure : vetopsy.fr d'après Fokin et Gautreau)
1. Le CPI de FAM21 du complexe WASH se lie à la protéine de coiffe CapZ de la dynactine pour mettre à jour le filament de dynactine, composé de 8 molécules d'Arp1 et d'une molécule d'actine β ( structure de la dynactine).
La dynactine et le motif CPI de FAM21 sont nécessaires à la génération de réseaux d'actine ramifiée endosomale.
Le filament de dynactine décoiffé est stable.
2. Le filament de dynactine peut ensuite être allongé avec de l'actine et fournit ainsi le premier filament d'actine pour la réaction de ramification d'Arp2/3 contrôlée par le domaine VCA/WCA de la sous-unité WASH.
- Il semblerait que la dynactine soit complexée avec la dynéine et ses adaptateurs de dynéine lorsqu'elle est intégrée dans le réseau d'actine ramifiée.
- Le filament peut accueillir des adaptateurs de la dynéine sur ses côtés et allonger un filament d'actine à partir de son extrémité barbée sans encombrement stérique (Cryo-EM shows how dynactin recruits two dyneins for faster movement 2018).
3. Le filament de type actine de la dynactine est intégré dans le réseau d'actine ramifié.
- Le tubule membranaire est allongé par la force de traction de la dynéine.
- WASH-Arp2/3 au niveau du col du tubule pousse contre la masse de l'endosome.
Ces forces antagonistes sont susceptibles d'étirer la membrane et de favoriser sa scission.
Endosomes de recyclage
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosquelettemicrofilaments d'actineFilaments intermédiairesMicrotubulesMatrice extracellulaireReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation