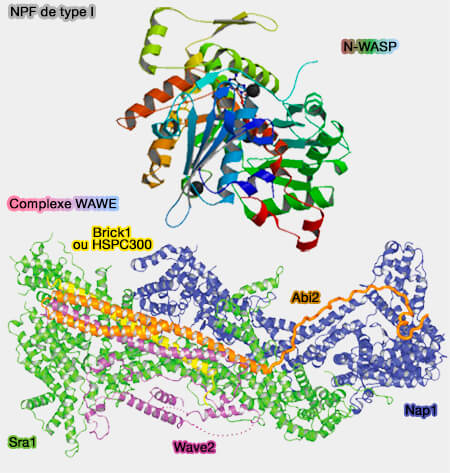
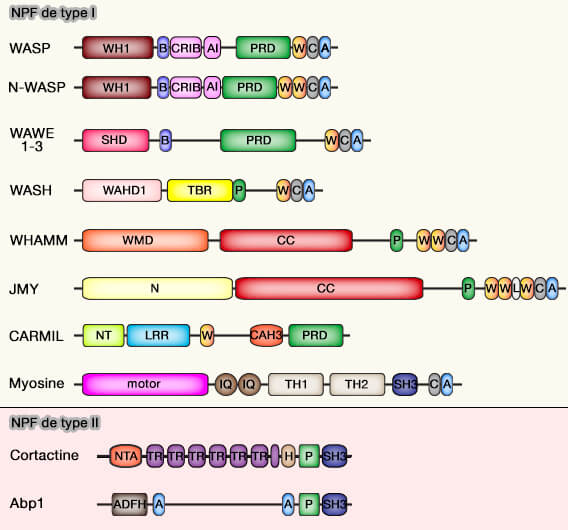
1. Les NPF de type I (famille WASP), possédant un domaine VCA, activent directement le complexe Arp2/3 en y provoquant un changement conformationnel et en se liant aux premiers monomères d'actine du nouveau filament.
2. Les NPF de type II, i.e. cortactine ou son homologue Abp1, HS1 (Haematopoietic cell-specific protein 1) ne possèdent pas de domaine VCA.
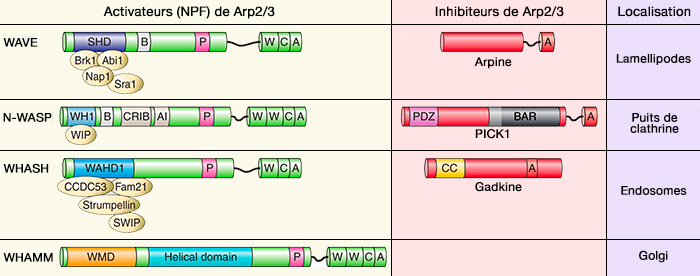
La famille WASP (Wiskott-Aldrich Syndrome Protein) contient quatre familles qui consistent à activer le complexe Arp2/3 à différentes localisations subcellulaires.
Par exemple, WAVE génère des réseaux d'actine ramifiés au niveau du cortex cellulaire et en particulier dans les saillies adhésives telles que les lamellipodes.
WASH génère des réseaux d'actine ramifiés à la surface des endosomes et autour des centrosomes.
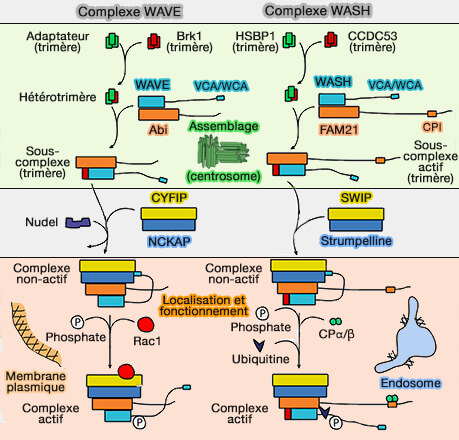
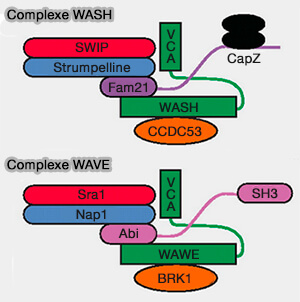
Les complexes multiprotéiques contenant des NPF sont responsables de la localisation subcellulaire du NPF, de son maintien dans un état inactif et de l'exposition au WCA lors de la liaison aux activateurs en amont (The Arp2/3 Regulatory System and Its Deregulation in Cancer 2018).
Organisation modulaire des protéines activatrices et inhibitrices d'Arp2 /3.
(Figure : vetopsy.fr d'après Molinie et Gautreau)
Exprimée dans les cellules hématopoïétiques, les mutations de N-WASP sont à l'origine du syndrome Wiskott-Aldrich, i.e. N-WASP présent dans les cellules n'est pas en mesure d'exécuter toutes les fonctions de WASP.
Une région interne unique de trente acides aminés (I30) dans WASP régule sa fonction dans la chimiotaxie des cellules T Jurkat.
WAWE1/2/3
1. Trois WAVE (WASP-family Verpolin homologous protein ou leur homologue SCAR) activent, entre autres, la formation des lamellipodes.




 protéines de stabilisation).
protéines de stabilisation).