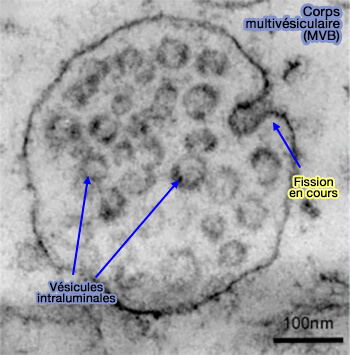
Les vésicules intraluminales (ILV), qui contiennent les cargos, i.e. en général les récepteurs membranaires quelquefois liés encore à leur ligand, se forment par invagination dans les endosomes et forment des endosomes multivésiculaires (MVE) ou corps multivésiculaires (MVB).
Vue d'ensemble des ILV et des MVE/MVB
Remarques essentielles
1. Les vésicules d'endocytose sont transportées vers les endosomes dans lesquels elles sont invaginées pour former des vésicules intraluminales (ILV).
Corps ou endosome multivésiculaire (MVB ou MVE)
(Figure : vetopsy.fr d'après Chivet et coll)
3. Les vésicules contenant ces ILV finissent par se séparer des régions tubulaires des endosomes précoces (qui bourgeonnent ensuite en vésicules séparées) et deviennent :
des vésicules porteuses endosomales (ECV ou Endosomal Carrier Vesicle),
et/ou des corps multivésiculaires (MVB ou multivesicular bodies ou MVE, endosomes multivésiculaires).
Quel que soit le devenir des ILV, dégradation, sécrétion ou récupération, on peut considérer ces vésicules, dont le pH est neutre lors de leur formation, comme des conteneurs unitaires emballant des quantités de cargaison dans la membrane ou dans la lumière.
4. Les ILV ou les MVE/MVB peuvent suivre :
un chemin principal, i.e. ils sont transportés vers les endosomes tardifs qui évoluent en endo-lysosomes pour atteindre les lysosomes pour la dégradation de leur contenu,
Distribution spatiale des endosomes multivésiculaires
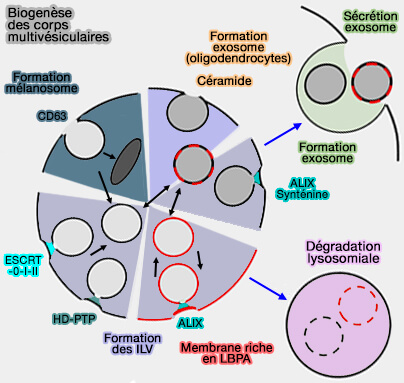
La formation de MVE/MVB commence probablement dans plusieurs emplacements subcellulaires pour séquestrer les cargos destinés à la dégradation ou à l'exocytose.
La distribution spatiale, la taille et le contenu des MVE/MVB ne sont pas uniformes et dépendent :
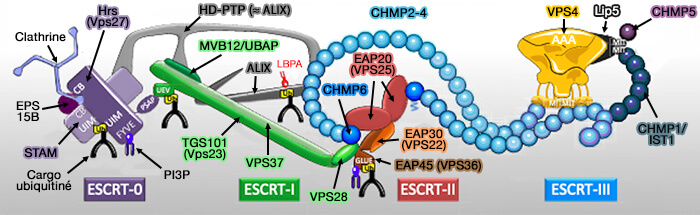
1. Les cargos sont chargés dans les ILV chez les eucaryotes par un ensemble de complexes protéiques appelé ESCRT (Endosomal Sorting Complex Required for Transport).
En l'absence d'ESCRT, les cargos transmembranaires sont incapables d'atteindre efficacement la lumière du lysosome.
2. Les complexes ESCRT ont un double rôle dans ce processus.
Ils induisent le bourgeonnement et la scission de la membrane endosomale loin du cytosol pour former des ILV contenant le cargo pour générer des MVE/MVB.
ESCRT-I, II et III provoquent la polymérisation du complexe ESCRT-III sous forme de filaments spiralés, et ainsi promeuvent l'invagination de la membrane contenant ces complexes.
ESCRT III termine la scission membranaire en utilisant l'AAA+ ATPaseVps4 pour la formation de la vésicule intraluminale.
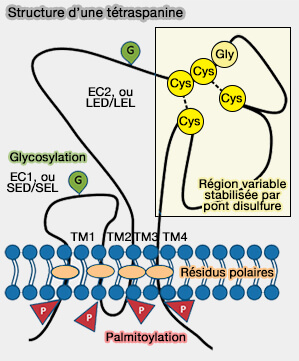
2. De nombreuses tétraspanines s'associent spécifiquement avec un nombre limité de protéines et d'autres tétraspanines pour former des microdomaineshautement organisés, constitués par des protéines d'adhésion, de signalisation et des adaptateurs, en particulier dans les systèmes immunitaire, reproducteur, génito-urinaire et auditif (Tetraspanins Function as Regulators of Cellular Signaling 2017).
3. Les tétraspanines sont impliquées dans la formation :
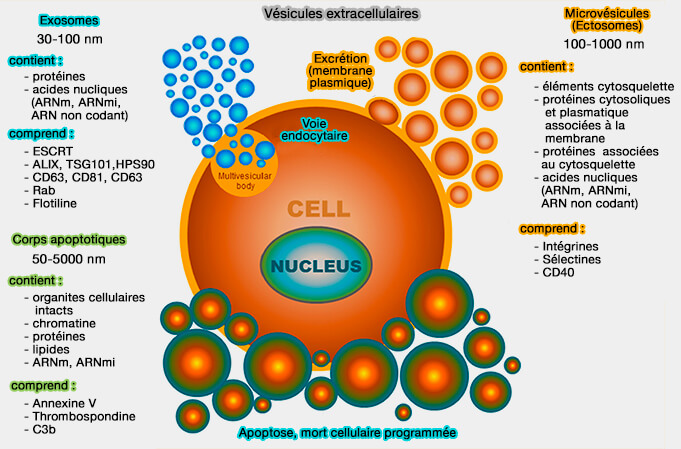
Vésicules extracellulaires
(Figure : vetopsy.fr d'après Jankovicova et coll)
4. Une voie dépendante d'ALIX et du complexe ESCRT favorise le tri et la livraison des tétraspanines aux exosomes (ALIX, ESCRT et exosomes).
Céramides
Les céramides sont essentiels pour la courbure de la membrane et le bourgeonnement des vésicules, aussi bien pour les MVE/MVB dans la voie indépendante du complexe ESCRT que pour les exosomes.
Le ciblage spécifique d’une protéine transmembranaire dans les ILV des MVE/MVB peut également se produire par le biais d’interactions protéine-protéine.
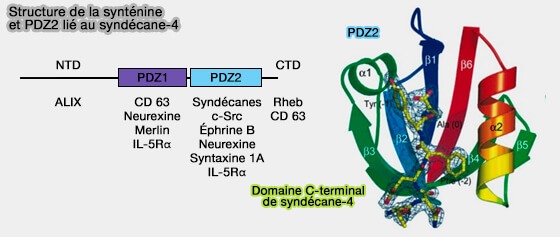
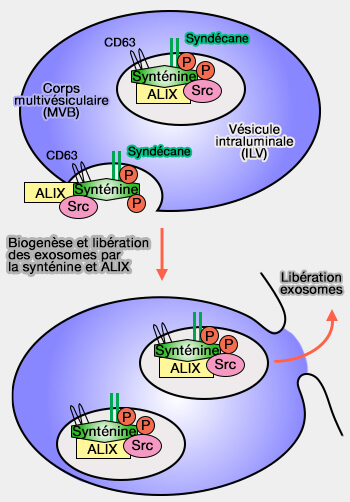
1. Les syndécanes, via leurs chaînes polysaccharidiques héparane sulfate, se lient à leur jonction cytoplasmique à la synténine pour participer à la formation des ILV.






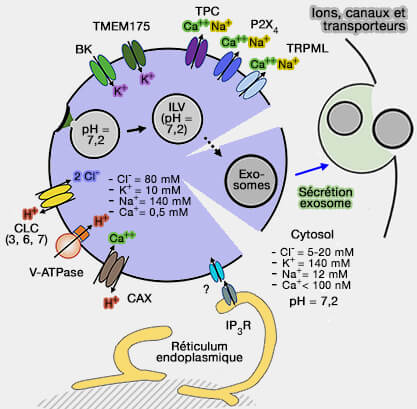
 fusion des lysosomes).
fusion des lysosomes).