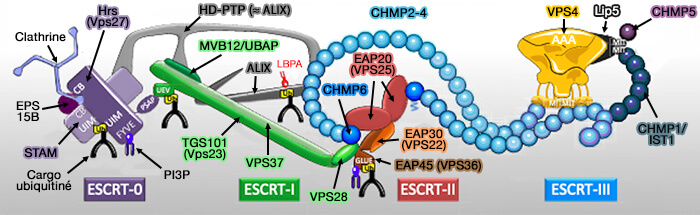
Le complexe ESCRT est composé de 5 complexes, dont ESCRT-III dont le mécanism qui entraîne le remodelage membranaire reste encore incertain.
Le complexe ESCRT (Endosomal Sorting Complex Required for Transport ou complexe de tri endosomal requis pour le transport) est composé de 5 complexes :
Une fois initiée, cette polymérisation de CHMP4 est favorisée par sa modification conformationnelle qui permet de recruter une succession de CHMP4 ( polymérisation des filaments).
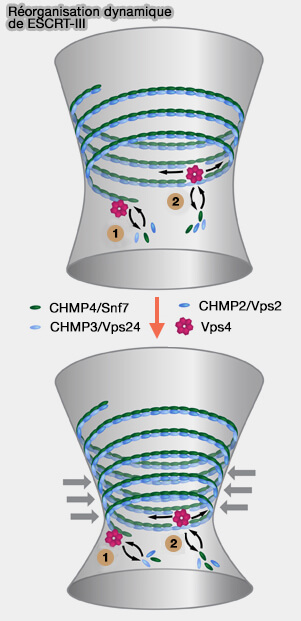
pour soutenir l'extension des filaments incurvés vers l'intérieur (1 sur la figure),
pour recruter Vps4 (AAA ATPase vacuolaire de tri) qui facilite le glissement des spires hélicoïdales voisines pour favoriser la constriction (2 sur la figure).
3. Vps4 induit le renouvellement des sous-unités au sein de l'ESCRT-III.
En effet, les polymères observés avec ces sous-unités (CHMP4/Snf7, ainsi que CHMP2/Vps2 et CHMP3/Vps4) ont un rayon supérieur à 15 nm, un peu plus petit avec Vps4, mais loin de la limite théorique de 3 nm requise pour la fission spontanée.
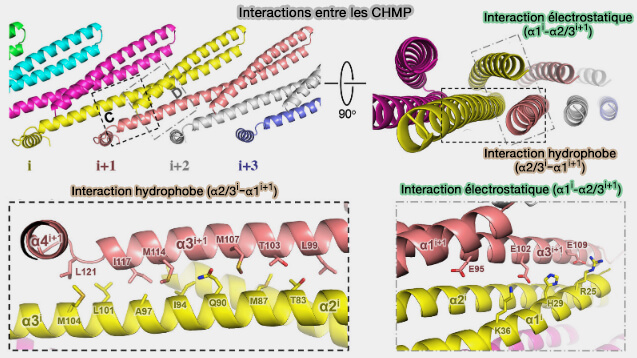
1. Dans la polymérisation des CHMP, le protomère (i), en conformation ouverte, interagit avec le protomère (i+1) à travers deux interfaces.
L'hélice étendue α2-α3 de (i) expose une surface hydrophobe sur α3 qui interagit avec α2-α3 de (i+1).
L'hélice α1 de (i) interagit électrostatiquement sur α3 qui interagit avec α2-α3 de (i+1).
Interactions des protéines CHMP ((CHMP4/Sndf7) et polymérisation
(Figure : vetopsy.fr d'après Tang
et coll)
Les interactions faibles entre filaments à simple brin ont une forte probabilité de rupture spontanée, ce qui explique pourquoi les microtubules et l'actine, par exemple, sont constitués de plusieurs protofilaments.
Toutefois, dans le cas des sous-unités ESCRT-III :
l'assemblage implique des interactions qui pourraient réduire considérablement le désassemblage spontané,
le processus d'activation nécessite une liaison membranaire, ce qui pourrait également réduire le désassemblage spontané.
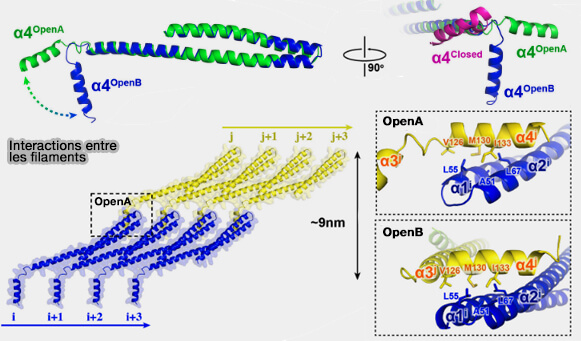
2. L'hélice α4 sert de liaison entre deux protofilaments différents (i et J), i.e. entre α4i et α1/2j.
Interactions des CHMP entre les filaments
(Figure : vetopsy.fr d'après Tang
et coll)
3. La polymérisation des ESCRT-III en monofilaments à simple brin est essentielle car elle implique :
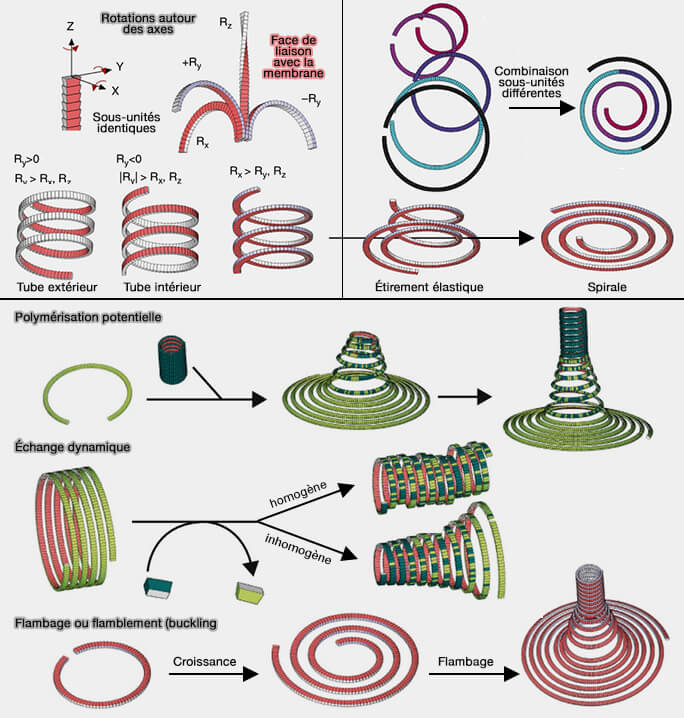
3. Des bobines et des cônes sont formés car les assemblages ESCRT tubulaires ne peuvent pas s'étendre indéfiniment lorsqu'ils sont entourés de lipides et doivent se fermer à une extrémité, comme c'est le cas du co-assemblage in vitro de CHMP2A et CHMP3 (Helical Structures of ESCRT-III are Disassembled by VPS4 2008), et bien d'autres…
4. Enfin, on trouve un mélange de toutes ces formes, i.e. spirales à tubes, cônes à tubes… ( cf. tableau).
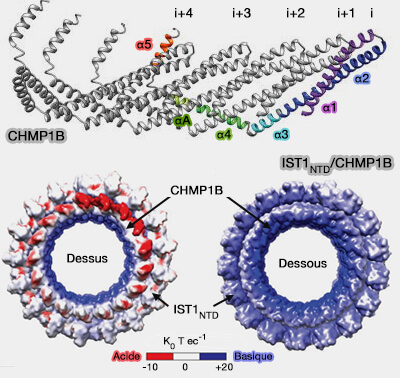
1. Chaque molécule de CHMP1B semble interagir avec quatre autres sous-unités CHMP1B qui se regroupent et traversent l'avant-bras de la sous-unité d'origine, permettant de grands changements de l'angle entre les monomères, et donc de courbure.
Dans les polymères plus rigides, comme les microtubules et l'actine, les interactions ne se trouvent qu'entre monomères voisins.
2. De plus, la main de l'hélice 5 saisit l'épaule de l'épingle à cheveux des quatre sous-unités, créant un contact analogue à l'interaction entre l'épingle à cheveux et l'hélice 5 dans les conformations ESCRT-III fermées.
Flexibilité des filaments
1. Normalement, les changements de courbure rompent facilement les interactions à courte distance, alors que dans le polymère CHMP1B, les interactions à longue distance médiées par une seule hélice α flexible, ne devraient pas être facilement rompues (Dynamic and elastic shape transitions in curved ESCRT-III filaments 2017).
Cela pourrait être à la base de la flexibilité inhabituellement grande des filaments CHMP4/Snf7 dont la longueur de rupture a été estimée entre 300 et 800 nm.
Cela pourrait également expliquer comment les structures ESCRT-III peuvent s'adapter à des courbures membranaires allant du micron à des dizaines de nanomètres de rayon, sans se casser.
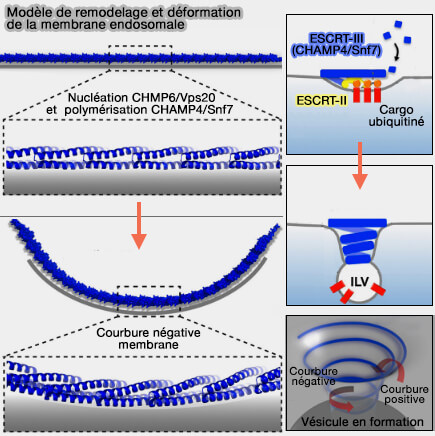
Remodelage et déformation de la membrane endosomale
(Figure : vetopsy.fr d'après Tang
et coll)
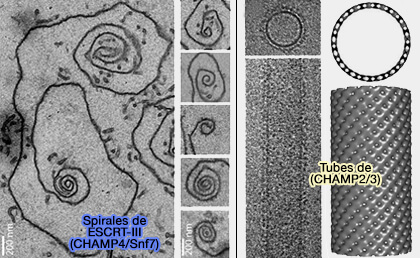
2. En effet, CHMP4/Snf7 forme des filaments très flexibles qui s'enroulent spontanément à la surface des membranes lipidiques.
La croissance d'un filament extensible en une spirale accumule une contrainte élastique, formant un cône terminé par une structure tubulaire.
Cette force, comme celle d'un ressort, déforme la membrane par compression latérale et est libérée par flambage ou flamblement, i.e. terme d'ingénierie des structures, buckling en anglais (Membrane buckling induced by curved filaments 2009).
Le flambage ou flambement est un phénomène d'instabilité d'une structure. Si cette structure est soumise à une charge progressivement croissante, lorsque la charge atteint un niveau critique, un élément peut soudainement changer de forme et on dit que la structure et le composant se sont déformés.




 polymérisation des filaments).
polymérisation des filaments).