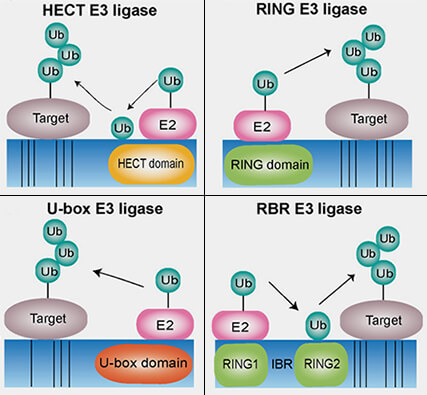
Les ligases E3 catalysent le transfert de l’ubiquitine sur des protéines spécifiques, garantissant la sélectivité du système ubiquitine-protéasome à travers des mécanismes enzymatiques et de reconnaissance des dégrons.
Elles comprennent plus de 600 membres chez l'homme, assurent la spécificité de la reconnaissance des substrats au sein du système ubiquitine-protéasome (UPS), marquant les protéines destinées à la dégradation ou à la régulation fonctionnelle.
Ces ligases réalisent le transfert direct de l’ubiquitine du complexe E2~Ub vers la protéine cible.
b. Les E3 ubiquitine ligases complexes peuvent être :
multiprotéiques, comme le complexe SCF (Skp1-Cullin-F-box) ou l'APC/C (Anaphase Promoting Complex/Cyclosome) par exemple, comprenant une seule sous-unité de recrutement de l’E2 et un ou plusieurs modules de reconnaissance du substrat,
oligomériques, souvent homodimériques ou hétérodimériques, qui peuvent contenir plusieurs domaines de recrutement pour E2 et/ou de reconnaissance du substrat, assurant à la fois la stabilité structurale et la régulation allostérique de l’activité enzymatique.
Mécanisme de l'ubiquitination et reconnaissance des substrats
Les ligases E3 assurent la spécificité du système ubiquitine-protéasome en catalysant le transfert de l’ubiquitine sur des protéines cibles précisément sélectionnées.
Familles des E3 ligases
(Figure : vetopsy.fr d'après Yang et coll)
Leur activité repose sur deux dimensions complémentaires.
1. Le mécanisme enzymatique est propre à chaque famille :
2. La reconnaissance sélective de signaux de dégradation appelés dégrons et des domaines de liaison à l’ubiquitine (UBD), détermine quelles protéines seront ubiquitinées.
Les dégrons sont étudiées dans des chapitres spécifiques.