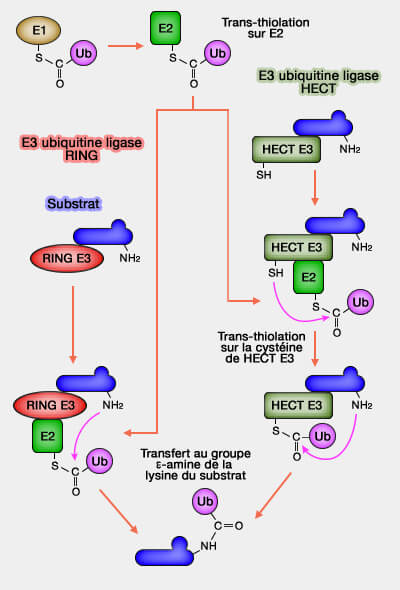
L’enzyme de conjugaison E2 reçoit l’ubiquitine activée par E1 et la transfère vers la ligase E3, assurant la continuité du cycle enzymatique d’ubiquitination.
L’ubiquitine doit, pour se fixer sur la protéine cible, processus appelé ubiquitination ou ubiquitinylation, recourir à une succession coordonnée d’enzymes spécifiques agissant de manière séquentielle.
L'enzyme d’activation (E1) active l’ubiquitine par une réaction ATP-dépendante pour former un intermédiaire thioester (E1~Ub).
L'enzyme de conjugaison (E2) reçoit ensuite l’ubiquitine activée depuis E1 pour générer le complexe E2~Ub.
L'enzyme ligase (E3) reçoit le complexe E2~Ub, reconnaît le substrat, oriente les partenaires et catalyse la formation de la liaison isopeptidique reliant l’ubiquitine à la protéine cible.
Ces trois étapes s’enchaînent de manière ordonnée pour aboutir à la formation d’un pont isopeptidique entre la glycine C-terminale (G76) de l’ubiquitine et une lysine du substrat, assurant ainsi la fixation covalente du signal ubiquitine.
Modèle du cycle catalytique canonique de E1
(Figure : vetopsy.fr d'après Straton et coll)
b. Ce transfert est précédé d’un changement conformationnel d’E1, rapprochant les cystéines catalytiques des deux enzymes, ce qui permet ( figure ci-dessus) :
la migration du thioester,
le relargage d’E1.
2. L’enzyme E2, désormais liée de manière transitoire à l’ubiquitine (complexe E2~Ub), collabore avec une E3-ubiquitine ligase (UBE3) pour :
3. On connait 11 E2 chez la levure et près d'une trentaine chez l'homme : plus d'une centaine ont été décrites (cf. Wikipedia) présentent une diversité fonctionnelle remarquable.
Certaines sont ubiquitaires, d’autres spécialisées dans des voies précises comme :
Conjugaison de la protéine UBL ATG8/LC3 à la
phosphatidyléthanolamine
Formation de la membrane autophagosomale.
BIRC6
BRUCE
—
E2/E3 hybride participant à la conjugaison d'UBL
et à la régulation de l'apoptose
UFC1
—
UFL1
E2 du système UFM1 (ubiquitin-like), impliquée dans
la traduction et la réponse au stress du réticulum endoplasmique
Structure des E2
Les enzymes E2 partagent une organisation structurale conservée centrée sur un domaine catalytique UBC (Ubiquitin-Conjugating), auquel peuvent s’ajouter des extensions N- ou C-terminales qui modulent leur spécificité et leurs interactions.
lors du transfert ultérieur de cette ubiquitine vers le substrat, en maintenant la réactivité du site catalytique et la conformation adéquate pour la formation de la liaison isopeptidique sous le contrôle de l’E3 ubiquitine ligase.
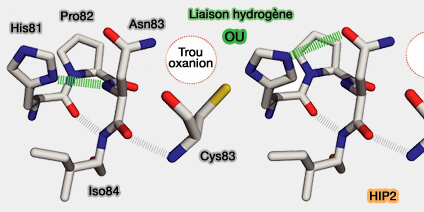
L’histidine contribue à stabiliser la conformation locale du site catalytique, tandis que l'asparagine (Asn) participe à la triade catalytique en modulant le microenvironnement acido-basique nécessaire à l’activation du substrat.
Par exemple, dans l’enzyme Ube2I, les résidus Asn85, Tyr87 et Asp127 jouent un rôle déterminant dans la réduction du pka de la lysine du substrat, facilitant ainsi l'attaque nucléophile sur la liaison thioester E2~Ub (ou E2~SUMO) et la formation de la liaison isopeptidique finale.
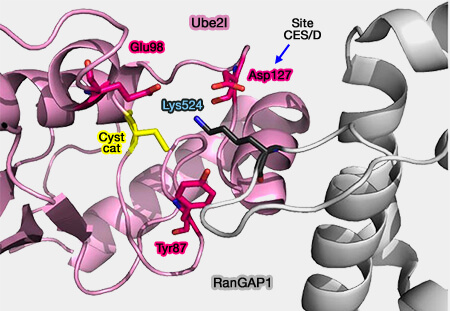
Coordination entre RanGAP1 et Ube21
(Figure : vetopsy.fr d'après Valimberti)
1. La taille et la charge négative de ce résidu, i.e. aspartate ou sérine phosphorylée par exemple dans Ube2a ou Rad6, permettent d’aligner la lysine du substrat avec la cystéine catalytique de l’E2, optimisant la géométrie du site actif et donc l’efficacité de l’ubiquitination.
Les E2 contenant une sérine peuvent ainsi être régulées par phosphorylation, introduisant un niveau de contrôle post-traductionnel supplémentaire.
b. Dans Ube2I, outre l’aspartate D127 du motif CES/D, d’autres résidus participent à la stabilisation du site catalytique.
L’acide glutamique 98 (Glu98) forme un trident électrostatique avec la lysine du substrat et D127,
La tyrosine 87 (Tyr87), par sa chaîne aromatique latérale, maintient la lysine dans la cavité catalytique et stabilise la transition menant à la formation de la liaison isopeptidique.
Interfaces d’interaction avec l’ubiquitine, E1 et E3
Les enzymes E2 assurent la coordination centrale entre l’activation de l’ubiquitine par E1 et son transfert final catalysé par les E3 ubiquitine ligases.
Elles interagissent successivement avec ces partenaires par l’intermédiaire de surfaces partiellement chevauchantes, ce qui impose un cycle précis de fixation et de libération.
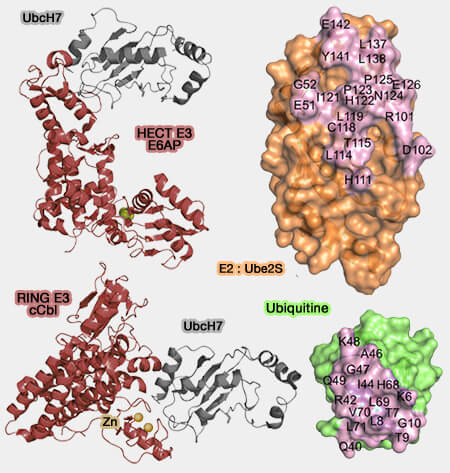
Liaisons des E2 avec l'ubiquitine et les E3
(Figure : vetopsy.fr d'après Lorenz et Wenzel)
Les interfaces d’interaction canoniques comprennent :
des résidus de l’hélice N-terminale (H1) et des boucles 4 et 7 (L4 et L7, parfois désignées L1 et L2), qui assurent les contacts principaux avec E1 et E3 ;
un site secondaire " backside ", opposé au site catalytique, pouvant fixer une ubiquitine non covalente ou un domaine UBD, jouant un rôle dans la régulation de la polyubiquitination ;
2. Ces variations topologiques expliquent la plasticité fonctionnelle des E2 et leur capacité à interagir avec des familles d’E3 très différentes, tout en conservant un cœur catalytique commun.
Les E2 possèdent des extensions N- et C-terminales spécifiques, qui modulent leurs propriétés structurales et leur spécificité fonctionnelle.
Ubiquitinations par les RING et les HECT
(Figure : vetopsy.fr)
1. Ces régions flexibles, parfois désordonnées, influencent leur stabilité, leur localisation subcellulaire et leur sélectivité d’interaction avec les E3 ubiquitine ligases.
Les E2 cytosoliques telles que UBE2D ou UBE2N interagissent avec plusieurs E3 de type RING, favorisant la formation de chaînes d’ubiquitine de topologies variées.
2. Ces extensions expliquent la grande diversité fonctionnelle des E2, plus de 600 isoformes ont été identifiées chez l’Homme, et leur adaptation à des voies ubiquitine-dépendantes spécifiques.
Ainsi, UBE2C/UbcH10, partenaire de l’APC/C (Anaphase-Promoting Complex/Cyclosome), illustre cette spécialisation.




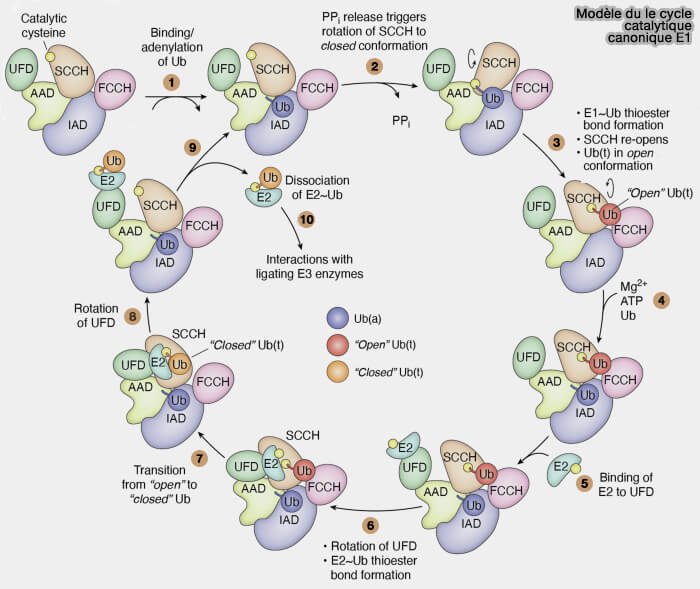
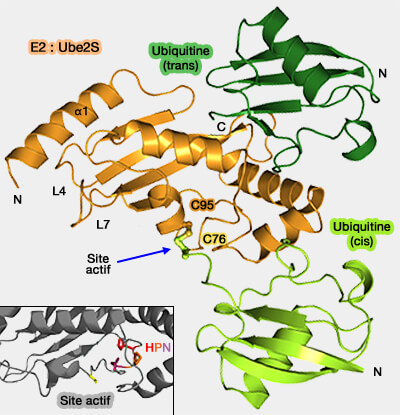
 figure ci-dessus) :
figure ci-dessus) :