Les dégrons terminaux sont des signaux moléculaires situés aux extrémités N- ou C-terminales des protéines, dont la nature chimique détermine leur stabilité en conditionnant leur reconnaissance par des ligases E3 spécifiques et leur dégradation ciblée par le protéasome via les voies N-end rule et C-end rule.
Les voies N-end rule et C-end rule constituent les exemples les plus emblématiques de dégrons terminaux, illustrant comment la nature chimique d’un simple acide aminé en position extrême, i.e. N- ou C-terminale, peut réguler avec une grande précision la stabilité, l’ubiquitination et le devenir d’une protéine.
Certaines protéases, comme les caspases ou les calpaïnes, exposent après clivage un nouveau résidu N-terminal reconnu par la voie Arg/N-end rule, reliant ainsi l’activation protéolytique à la dégradation sélective des substrats.
La voie N-end rule représente un modèle canonique de dégradation dépendante des dégrons, fondé sur la reconnaissance de l’acide aminé situé à l’extrémité N-terminale comme déterminant majeur de la demi-vie protéique en déclenchant son ubiquitination et sa dégradation ciblée par le protéasome.
1. Le principal déterminant d'un N-dégron est un résidu soit modifié, soit non modifié.
2. Un pro-N-dégron (précurseur d'un N-dégron) est une séquence spécifique ou une conformation d'une chaîne de polypeptide qui peut être clivée ou modifiée pour produire un résidu N-terminal déstabilisant. Pour cela, les résidus sont classés en :
Ndp pour primaires : résidu N-terminal déstabilisant ou résidu N-terminal non modifié directement reconnu par une N-recognin,
Ndt pour tertiaires : résidu dont la déstabilisation nécessite deux modifications préliminaires.
Branches de la règle du N-terminal
Chez les Eucaryotes, la voie de la règle N-terminale se compose de deux branches.
Ac/règle N-terminale (Ac/N-end rule)
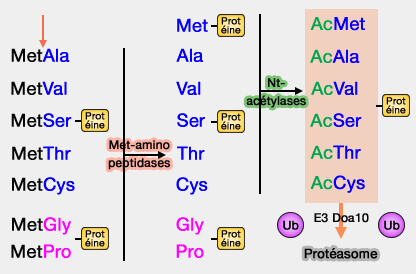
1. L'Ac/règle N-terminale (Ac/N-end rule), présente aussi chez les Procaryotes, cible les protéines contenant les résidus Nα-terminaux acétylés, qui sont des Ac N-dégrons, comme :
Ac/N-end rule
(Figure : vetopsy.fr)
de petits résidus non chargés comme l'alanine (A), la valine (V), la sérine (S), la thréonine (T) ou la cystéine (C).
Ce sont donc des Nds (secondaires), car ils doivent être acétylés pour être reconnus par l'E3 ubiquitine ligases.
b. Il semble que cette Nt-acétylation soit irréversible, contrairement aux autres acétylations internes dans lesquelles des déacétylases peuvent inverser le processus.
Certains résidus sont rarement Nt-acétylés comme la glycine (G) ou la proline (P).
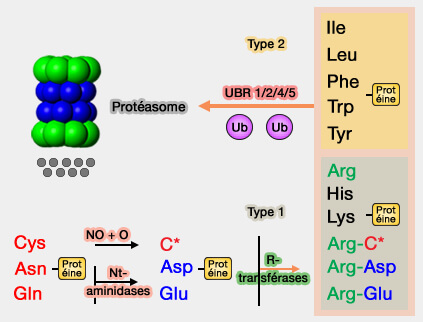
L'asparagine et la glutamine doivent d'abord être converties par une Nt-aminidase en Asp et Glu, et la cystéine oxydée en présence de NO et d'oxygène, avant d'être arginylées comme précédemment: ce sont donc des Ndt (tertiaires).
Cette règle N-terminale n'implique pas l'ubiquitination chez les Procaryotes.
3. Les N-recognins (N-recognin) sont des ligases E3 spécialisées, i.e. UBR 1/2/4/5, qui détectent et lient sélectivement les N-dégrons via des domaines fonctionnels :
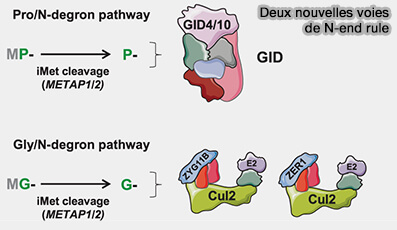
La Pro/N-end rule et la Gly/N-end rule, de découverte récente, sont des sous-voies distinctes de la voie N-end rule, qui fonctionnent sur le même principe général, i.e. la nature de l’acide aminé N-terminal détermine la stabilité de la protéine.
Elle est reconnue par des ligases E3 spécifiques comme
Cette voie intervient notamment dans la surveillance de protéines mal adressées aux mitochondries ou au réticulum endoplasmique.
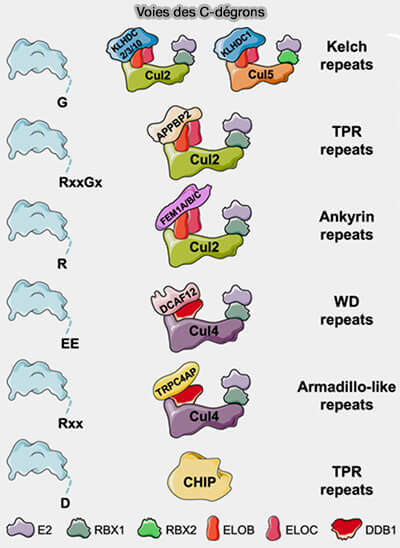
Dégrons C-terminaux et voie C-end rule
La C-end rule (ou voie C-dégron) repose sur le même principe conceptuel que la N-end rule, mais joue un rôle plus spécialisé et moins étendu dans la régulation protéique.