Les E3 ubiquitine ligases reconnaissent les substrats et catalysent le transfert final de l’ubiquitine, conférant la spécificité au système ubiquitine-protéasome.
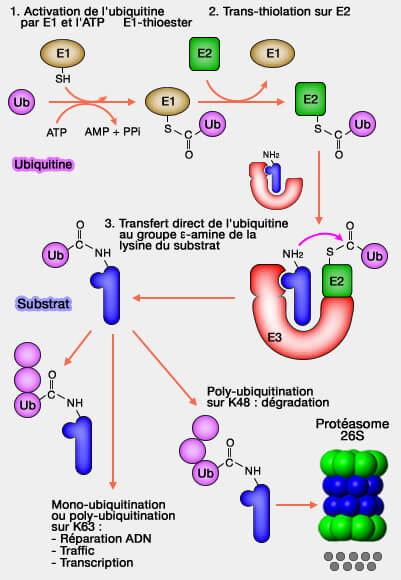
L’ubiquitine doit, pour se fixer sur la protéine cible, processus appelé ubiquitination ou ubiquitinylation, recourir à une succession coordonnée d’enzymes spécifiques agissant de manière séquentielle.
L'enzyme d’activation (E1) active l’ubiquitine par une réaction ATP-dépendante pour former un intermédiare thioester (E1~Ub)
L'enzyme de conjugaison (E2) reçoit ensuite l’ubiquitine activée depuis E1 pour générer le complexe E2~Ub.
L'enzyme ligase (E3)reçoit le complexe E2~Ub, reconnaît le substrat, oriente les partenaires et catalyse la formation de la liaison isopeptidique reliant l’ubiquitine à la protéine cible.
Ces trois étapes s’enchaînent de manière ordonnée pour aboutir à la formation d’un pont isopeptidique entre la glycine C-terminale (G76) de l’ubiquitine et une lysine du substrat, assurant ainsi la fixation covalente du signal ubiquitine.
indirectement, par l’intermédiaire d’un thioester transitoire formé sur la cystéine catalytique de l’E3, caractéristique des familles HECT et RBR.
1. L’activité de l’enzyme E3 ne se limite pas à transférer mécaniquement l’ubiquitine : elle consiste avant tout à organiser la configuration catalytiquement réactive entre les partenaires du complexe.
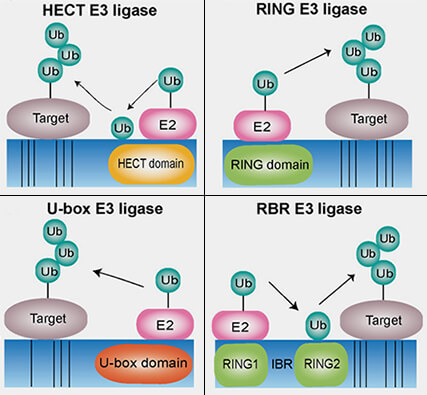
Familles des E3 ligases
(Figure : vetopsy.fr d'après Yang et coll)
L’E3 induit une déformation conformationnelle du complexe E2~Ub, en particulier au niveau de la liaison thioester entre E2 et l’ubiquitine.
Cette contrainte structurelle place l’ubiquitine dans un état " tendu " ou activé, favorable à la rupture contrôlée de la liaison thioester et au transfert du groupement carbonyle terminal vers le substrat.
b. E3 oriente les groupements réactifs.
En rapprochant spatialement l’ubiquitine donneuse et la lysine (ou l’amine N-terminale) du substrat, l’E3 aligne les orbitales électroniques du groupement carbonyle de la Gly76 de l’ubiquitine et de l’amine ε de la lysine cible.
Cette orientation optimale facilite la formation de la liaison isopeptidique, tout en limitant les réactions parasites.
c. E3 abaisse l’énergie d’activation de la réaction de transestérification ou d’aminolyse.
L’E3 stabilise l’état de transition de la réaction, réduisant ainsi l’énergie nécessaire à la transestérification dans le cas des E3 de type HECT et RBR, ou à l’aminolyse directe, i.e. attaque nucléophile d’un groupement carbonyle par une amine, pour les E3 de type RING.
Ce rôle catalytique subtil explique que certaines mutations de résidus de surface des E3, sans affecter la chimie du site actif, puissent abolir l’activité enzymatique en perturbant cet équilibre conformationnel de pré-organisation.
2. La nature de la liaison formée dépend du site accepteur :
plus rarement,sérine, thréonine ou cystéine dans des ubiquitinations non canoniques, comme celle des protéines NEMO ou MAVS, observée au cours des réponses antivirales ou inflammatoires.
Ces variations dans le site accepteur et la nature du lien formé reflètent la diversité fonctionnelle des ligases E3, dont le mécanisme catalytique diffère selon la famille structurale à laquelle elles appartiennent.
Variantes du mécanisme catalytique selon les famillesd
Le mécanisme catalytique des ligases E3 présente deux variantes principales, selon que le transfert de l’ubiquitine s’effectue directement ou via un intermédiaire covalent.
Cette distinction reflète la nature chimique du rôle joué par l’E3 dans la réaction et la diversité structurale de ses domaines catalytiques.
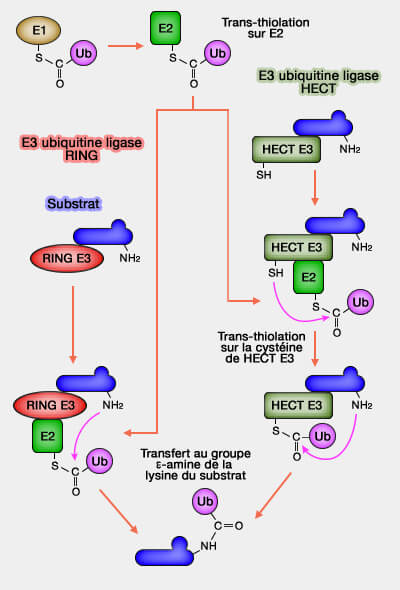
Transfert direct par les familles RING et U-box
Les ligases E3 des familles RING (Really Interesting New Gene) et U-box catalysent un transfert direct de l’ubiquitine depuis l’enzyme de conjugaison E2~Ub vers la protéine cible, sans formation d’intermédiaire covalent sur l’E3.
Ubiquitination par la famille RING
(Figure : vetopsy.fr)
1. Le domaine RING, stabilisé par deux ions Zn++, et le domaine U-box, de repli similaire, mais dépourvu de zinc, jouent un rôle analogue en servent de plateformes de positionnement permettant la liaison simultanée du complexe E2~Ub et du substrat :
une face interagit avec l’enzyme E2 chargée d’ubiquitine (E2~Ub),
l’autre reconnaît la protéine substrat directement ou via une sous-unité adaptatrice.
a. Le rôle catalytique principal du domaine RING consiste à activer le complexe E2~Ub en favorisant sa transition vers une conformation " fermée " ou tendue.
Dans cet état, l’ubiquitine est partiellement détachée de l’E2 et orientée de façon optimale pour le transfert de son groupement carbonyle terminal vers la lysine du substrat.
Cette activation résulte d’interactions précises entre la surface hydrophobe de l’ubiquitine et des résidus conservés du domaine RING, souvent Phe, Leu ou Ile, qui stabilisent l’état de transition du lien thioester E2~Ub.
b. La réaction procède par une aminolyse directe, i.e. le groupement amine ε de la lysine du substrat attaque le carbone carbonyle du lien thioester E2~Ub, entraînant la formation de la liaison isopeptidique entre la Gly76 de l’ubiquitine et la lysine cible.
L’E3 n’intervient pas chimiquement, mais elle oriente les partenaires et stabilise l’état de transition.
2. De nombreuses E3 de type RING fonctionnent dans des complexes modulaires, où la reconnaissance du substrat est assurée par des sous-unités distinctes :
les CRL (Cullin-RING Ligases), dont la sous-unité Rbx1/Rbx2 porte le domaine RING et recrute l’E2~Ub, tandis que les adaptateurs, i.e. F-box, BTB, DCAF assurent la spécificité du substrat.
Ubiquitinations par les RING et les HECT
(Figure : vetopsy.fr)
Ce mode d’organisation confère aux E3 RING une grande rapidité catalytique et une spécificité élevée, adaptées à la régulation fine de processus tels que :
La famille HECT et la famille RBR effectuent ce processus en deux étapes successives, impliquant la formation d’un intermédiaire thioester sur la cystéine catalytique de l’E3.
L'ubiquitine est transférée du complexe E2~Ub vers une cystéine catalytique de l’E3, formant un intermédiaire covalent E3~Ub.
L’ubiquitine est ensuite transférée de cette cystéine vers la lysine ou parfois vers un autre acide aminé du substrat cible, produisant la liaison isopeptidique définitive.
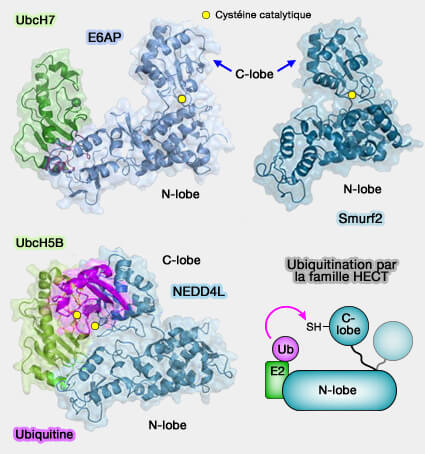
Ce mécanisme confère à ces ligases une plus grande plasticité catalytique, leur permettant de contrôler finement la topologie et la longueur des chaînes d’ubiquitine formées.
1. Les ligases de type HECT et RBR sont plus lentes que les RING, mais leur capacité à former un intermédiaire covalent leur confère une souplesse catalytique supérieure.
Elles peuvent :
reconfigurer la topologie des chaînes ubiquitine,
moduler la sélectivité des substrats,
intégrer des mécanismes de régulation allostérique ou post-traductionnelle, i.e. phosphorylation, ubiquitination secondaire, interaction cofacteur.
2. Ce mode de transfert indirect permet un contrôle fin et modulable de l’ubiquitination, essentiel dans des processus tels que :





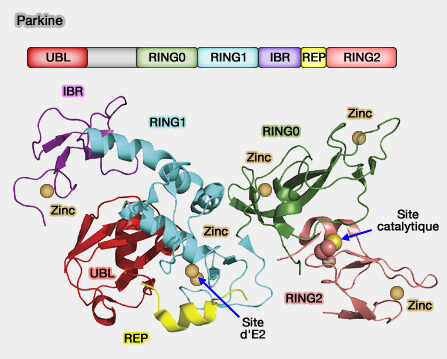
 structure des RBR) :
structure des RBR) :