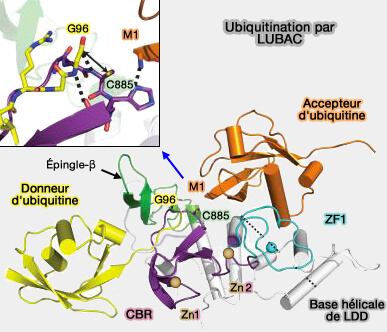
Les ligases E3 de type RBR combinent les mécanismes RING et HECT en formant un intermédiaire thioester avant de transférer l’ubiquitine sur leurs substrats.
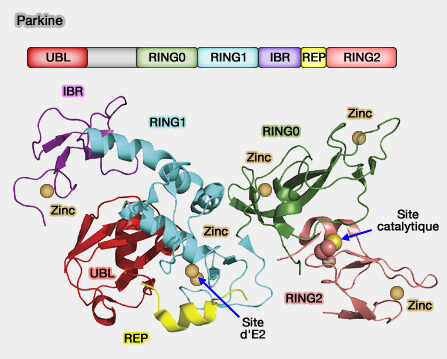
un domaine IBR (In-Between-RING) central, de structure apparentée à RING2,
Parkine de la famille des E3 ubiquitine ligases RBR
(Figure : vetopsy.fr d'après Berndsen)un domaine RING2 C-terminal, responsable de l’activité catalytique et fonctionnant selon un mécanisme de type HECT plutôt que RING.
Remarque : le domaine RBR est constitué de trois modules structuraux successifs RING1, IBR et RING2.
2. Dans sa forme compacte inactive, la parkine est auto-inhibée.
un motif zinc-finger (ZF2) dans le domaine hélical C-terminal du domaine LDD.
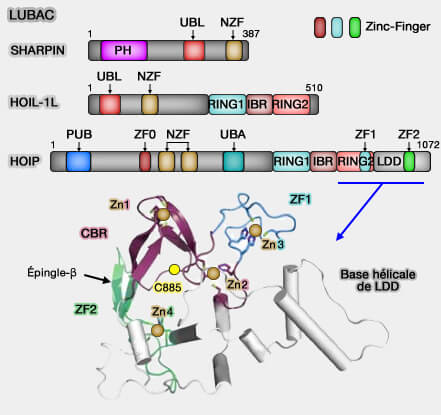
3. Le domaine LDD (Linear ubiquitin chain Determining Domain) de HOIP constitue l’élément catalytique terminal du complexe LUBAC, essentiel à la formation des chaînes linéaires d’ubiquitine (Structural basis for ubiquitylation by HOIL-1 2023).
a. Le LDD assure la reconnaissance et la stabilisation du couple d’ubiquitines impliqué dans la réaction.
L’ubiquitine donneuse est liée de manière covalente à la cystéine catalytique (Cys885) du domaine RING2 de HOIP.
L’ubiquitine accepteur est recrutée de manière non covalente par le LDD, via une interaction structurée autour de son extrémité N-terminale (Met1).
b. Le domaine LDD adopte une conformation hélicoïdale allongée, formant une interface double :
une interface avec RING2 qui aligne la Cys885 et le groupement carbonyle de Gly76 de l’ubiquitine donneuse,
une interface avec l’ubiquitine accepteur, où la His887 du LDD joue un rôle clé dans la catalyse.
c. His887 agit comme une base générale, déprotonant l’amine N-terminale de la Met1 de l’ubiquitine accepteur et l'activant pour détruire la liaison thioester E2~Ub.
Ce processus active l’amine pour une attaque nucléophile sur le groupement carbonyle du thioester RING2~Ub, ce qui conduit à la formation d’une liaison peptidique linéaire entre les deux ubiquitines.
d. En imposant cette géométrie stricte entre les ubiquitines, le LDD exclut la participation des lysines de l’ubiquitine accepteur, empêchant ainsi la formation de chaînes K48 ou K63, et déterminant donc la linéarité spécifique des chaînes M1, caractéristique unique du LUBAC.
4. Les chaînes M1-ubiquitine ainsi formées jouent un rôle fondamental dans :





 complexe E2~ubiquitine).
complexe E2~ubiquitine).