


Cycle cellulaire
Reproduction cellulaire : mitose
Anaphase
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

L’anaphase, qui dans la mitose suit la métaphase, débute par la séparation des chromatides sœurs et leur migration symétrique vers les deux pôles du fuseau.
L’anaphase correspond à la phase où la rupture de la cohésion centromérique libère les chromatides sœurs et déclenche leur migration polarisée, portée par la dynamique coordonnée des microtubules du fuseau mitotique.
Cette séparation s’effectue en deux mouvements distincts, allongement des microtubules polaires et traction sur les microtubules kinétochoriens, qui garantissent un partage équitable du matériel génétique entre les futures cellules filles.
Événements cellulaires de l'anaphase :
ségrégation chromosomique
La ségrégation chromosomique correspond à l’ensemble des événements de l’anaphase combinant :
- l’anaphase A, qui assure la séparation et le déplacement des chromatides sœurs vers les pôles par le raccourcissement des microtubules kinétochoriens,
- l’anaphase B, qui augmente la distance entre les pôles du fuseau par son allongement, renforçant la séparation spatiale des lots chromosomiques.
Séparation des chromatides sœurs
La séparation des chromatides sœurs est déclenchée par l’activation de l’APC/CCdc20, ce qui permet leur individualisation immédiate et le début de leur migration vers les pôles, par :
- la dégradation de la sécurine,
- la libération de la séparase,
- le clivage de la cohésine, qui provoque la rupture de la cohésion centromérique.
Migration des chromatides soeurs
La séparation des chromatides sœurs s’accompagne de deux mouvements mécaniques complémentaires et le plus souvent concomitants, qui assurent leur migration coordonnée vers les pôles du fuseau.
Anaphase A : raccourcissement des microtubules kinétochoriens
Lors de l'anaphase A, le raccourcissement des microtubules kinétochoriens assure la traction vers les pôles.
1. Les microtubules kinétochoriens se dépolymérisent à leur extrémité (+), générant une force dirigée vers les centrosomes.
a. Les microtubules kinétochoriens sont ancrés au kinétochore par leur extrémité (+) via le réseau d’interface constitué principalement par le complexe Ndc80 et ses partenaires ( complexe KMN).
complexe KMN).
- La dépolymérisation de cette extrémité, initiée par la perte de la coiffe GTP, et modulée par des enzymes dépolymérisantes telles que les kinésines-13 (Kif2/MCAK), entraîne le recourbement des protofilaments de tubuline.
- Cette courbure est mécaniquement couplée au kinétochore par le complexe Ndc80 et par le complexe Ska (Ska1-Ska2-Ska3), qui maintiennent l’attachement tout en autorisant le glissement contrôlé.
b. L’énergie élastique libérée par le recourbement des protofilaments est ainsi convertie en force de traction, le kinétochore, restant couplé à l’extrémité en rétraction, est entraîné vers l’extrémité (-) du microtubule, ancrée au centrosome, assurant la traction des chromatides vers les pôles.
2. Les moteurs mitotiques jouent ici un rôle auxiliaire assuré principalement par la dynéine et, de façon plus marginale, par certaines kinésines kinétochoriennes comme CENP-E, contribuant à l’ajustement et à la régularité du déplacement des chromatides.
Anaphase B : écartement des pôles et allongement du fuseau
En anaphase B, l’écartement des pôles du fuseau et l'allongement du fuseau résultent de la combinaison ( fuseau en anaphase) :
- de forces d’expansion générées dans la zone médiane par le glissement des microtubules polaires antiparallèles
- de forces de traction exercées par la dynéine corticale sur les microtubules astraux.
1. Dans la région médiane du fuseau mitotique, les microtubules polaires issus des deux centrosomes s’interpénètrent et s’organisent en faisceaux antiparallèles, i.e. orientés tête-bêche avec leurs extrémités (+) dirigées l’une vers l’autre.
Cette organisation antiparallèle définit la zone médiane (spindle midzone), zone fonctionnelle distincte de la plaque équatoriale car elle ne porte pas les chromosomes, mais constitue un module mécanique dédié à l’allongement du fuseau.
a. Le glissement et la dynamique des microtubules polaires dans cette zone médiane entraînent un déplacement progressif des pôles du fuseau et participent à l’allongement global du fuseau mitotique.
- Bien que les microtubules polaires n’interagissent pas directement avec les chromosomes, leur organisation en faisceaux antiparallèles et leur glissement relatif entraînent un déplacement progressif des pôles du fuseau.
- Les chromatides sœurs, restant attachées à ces pôles via les microtubules kinétochoriens, sont ainsi entraînées passivement, ce qui augmente la distance qui les sépare.
b. L’écartement des pôles du fuseau permet d’amplifier et de sécuriser la ségrégation chromosomique après leur séparation initiale, en augmentant la distance entre les deux masses chromosomiques.
- Il complète l’anaphase A, limite les interactions résiduelles entre masses chromosomiques, crée un espace central favorable à l’organisation de la zone médiane et prépare la formation ultérieure du corps intermédiaire.
- Il rend enfin la séparation des chromatides mécaniquement irréversible avant l’entrée en télophase.
2. En parallèle du glissement des microtubules polaires dans la zone médiane, des forces de traction exercées sur les microtubules astraux par la dynéine du cortex cellulaire contribuent au déplacement des pôles du fuseau.
Organisation de la zone médiane
En anaphase B, l’allongement contrôlé du fuseau repose sur l’organisation d’une zone médiane structurée, capable de maintenir un chevauchement antiparallèle stable entre microtubules polaires tout en autorisant leur glissement sous l’effet des forces mécaniques (Geometry of antiparallel microtubule bundles regulates relative sliding and stalling by PRC1 and Kif4A 2018).
1. PRC1 (Protein Regulator of Cytokinesis 1) assure la cohésion structurale de la zone médiane du fuseau en reconnaissant et en stabilisant le chevauchement antiparallèle des microtubules polaires (faisceautage).
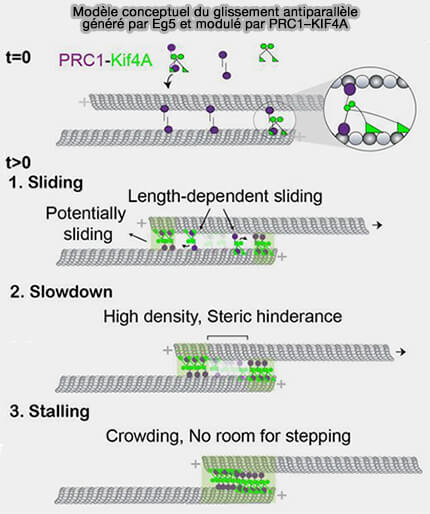
généré par Eg5 et modulé par PRC1-KIF4A
(Figure : vetopsy.fr d'après Wijeratne et Subramanian)
a. Dès la prométaphase et la métaphase, PRC1 assure une cohésion locale du chevauchement sans permettre son extension, mais agit comme un facteur de stabilisation et de confinement mécanique.
La zone médiane est courte et contrainte, les faisceaux antiparallèles ne forment un rail long et continu compatible avec un allongement net du fuseau malgré la présence de moteurs d’expansion.
b. En anaphase B, lorsque le glissement interpolaire devient productif, PRC1 assure la cohésion structurale du chevauchement sous l’effet des forces d’expansion.
- En maintenant l’alignement et la continuité du chevauchement sous l’effet des forces d’expansion, PRC1 permet un glissement efficace et spatialement confiné des microtubules lors de l’anaphase B.
- Elle définit ainsi une plateforme médiane stable, indispensable à la coordination des moteurs et régulateurs de la zone médiane et à la transition progressive du fuseau vers l’organisation du midbody en télophase.
2. KIF4A n’agit pas comme moteur d’écartement, mais comme régulateur de l’architecture de la zone médiane en limitant l’allongement excessif des microtubules dans la zone de chevauchement (Geometry of antiparallel microtubule bundles regulates relative sliding and stalling by PRC1 and Kif4A 2018)
a. Son recrutement spécifique dans la zone médiane est fortement dépendant de PRC1, par une interaction protéine-protéine directe, ce qui positionne KIF4A au site précis où la croissance des microtubules polaires antiparallèles doit être régulée.
b. Une fois recrutée par PRC1, KIF4A se déplace le long des microtubules et s’accumule préférentiellement vers l’extrémité (+) des microtubules chevauchants, se positionnant ainsi exactement au site de la polymérisation.
- À cet endroit, KIF4A réduit l’accessibilité des sites d’incorporation des dimères d'αβ-tubuline, ralentissant localement la vitesse d’ajout de nouvelles sous-unités.
- La croissance des microtubules est ainsi freinée spécifiquement dans la zone de chevauchement, sans affecter la dynamique globale du fuseau.
c. En l’absence de KIF4A, les microtubules antiparallèles s’allongent excessivement, la zone médiane devient trop large et instable, le chevauchement est mal défini et le glissement assuré par Eg5/KIF11 devient inefficace ou désorganisé.
3. Les kinésines-6, principalement MKLP2/KIF20A et MKLP1/KIF23, interviennent dans l’organisation et la cohésion des faisceaux antiparallèles de la zone médiane au cours de l’anaphase B.
a. Elles ne constituent pas le moteur principal de l’écartement des pôles, mais interviennent dans la structuration et la stabilisation du chevauchement antiparallèle sur lequel s’exercent les forces d’expansion.
- MKLP2/KIF20A s’accumule précocement sur les microtubules antiparallèles de la zone médiane et crée l’environnement structural permettant le recrutement des complexes tardifs, dont MKLP1.
- MKLP1/KIF23 s’accumule secondairement via sa participation au complexe centralspindlin, lorsque la zone médiane est déjà organisée, et assure la cohésion et la stabilisation finales nécessaires à la formation du midbody en télophase.
b. Les kinésines-6 assurent une fonction d’intégration structurelle, en maintenant l’alignement et la continuité des microtubules antiparallèles, condition nécessaire pour que le glissement généré par Eg5/KIF11, stabilisé par KIF15 et contrôlé localement par KIF4A, s’exerce de manière efficace et spatialement contrôlée.
4. La géométrie du chevauchement antiparallèle définie par PRC1 et KIF4A conditionne directement l’efficacité des forces d’expansion ultérieures.
En limitant l’allongement excessif du chevauchement antiparallèle, KIF4A empêche la dispersion des forces de glissement sur une zone trop étendue, ce qui permet de concentrer l’action d’Eg5, d’éviter les glissements inefficaces et de maintenir un allongement interpolaire directionnel et mécaniquement stable.
Forces mécaniques du fuseau en anaphase B
Forces internes au fuseau central
Forces d’expansion interne au fuseau central
En anaphase B, l’écartement des pôles du fuseau est assuré par des forces d’expansion générées dans la zone médiane par le glissement des microtubules polaires antiparallèles.
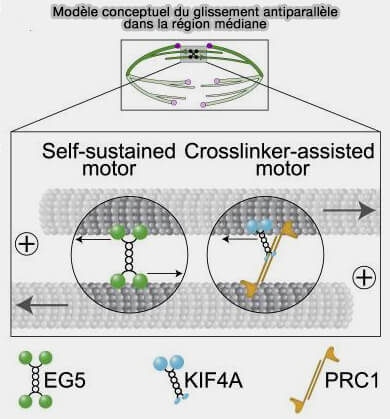
(Figure : vetopsy.fr d'après Wijeratne et Subramanian)
1. Dans cette zone de recouvrement, Eg5/KIF11 est la kinésine dominante (Measuring Pushing and Braking Forces Generated by Ensembles of Kinesin-5 Crosslinking Two Microtubules 2015).
a. Elle est tétramérique et possède deux domaines moteurs à chaque extrémité ((Microtubule-sliding modules based on kinesins EG5 and PRC1-dependent KIF4A drive human spindle elongation 2021)
- Elle se lie, en même temps, à deux microtubules orientés en sens inverse et les fait glisser l’un par rapport à l’autre.
- Elle génère ainsi une force d’écartement des pôles et provoque l’allongement progressif du fuseau caractéristique de l’anaphase B.
b. Toutefois, le glissement assuré par Eg5/KIF11 s’exerce dans un contexte hautement dynamique, marqué par le renouvellement continu des microtubules, la variabilité des charges mécaniques et la nature transitoire des interactions moteur-microtubule.
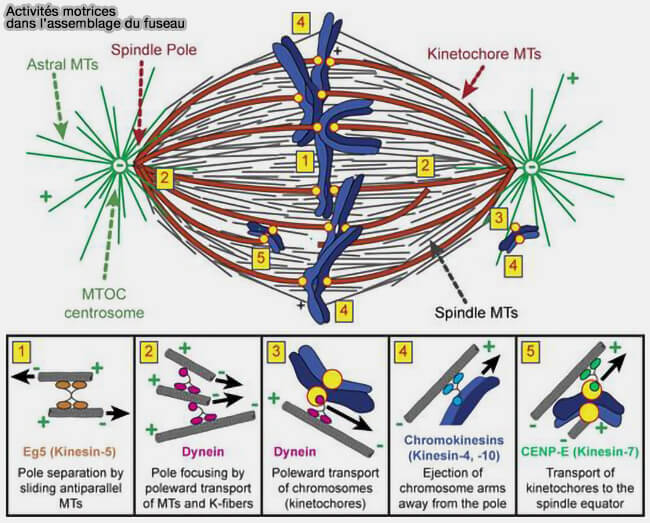
(Figure : vetopsy.fr d'après Heald et Khodjakov)
2. En conséquence, KIF15 agit comme un renfort mécanique du glissement antiparallèle généré par Eg5/KIF11, en empêchant la dissociation des microtubules antiparallèles sous l’effet des forces d’écartement.
KIF15 se positionne dans la zone de chevauchement en se liant simultanément à deux microtubules opposés, où elle agit comme un renfort mécanique et un élément porteur du système de glissement.
- Son activité motrice est faible, mais sa capacité à rester attachée sous charge est élevée, de sorte qu’une partie de la tension favorisant le détachement d’Eg5/KIF11 est absorbée par KIF15.
- Grâce à ce partage de charge, le chevauchement antiparallèle ne s’effondre pas, la distance entre pôles est maintenue, les microtubules restent correctement orientés.
- La géométrie bipolaire est conservée, même si Eg5 est moins active ou si la dynamique des microtubules est élevée.

Eg5/KIF11 est le moteur qui fait avancer le système, KIF15 est le moteur qui empêche le système de reculer ou de décrocher, i.e. il ne crée pas la séparation mais en empêche la perte.
Forces antagonistes limitant l’écartement des pôles
L’écartement des pôles du fuseau en anaphase B résulte d’un équilibre dynamique entre forces d’expansion et forces antagonistes, garantissant un allongement contrôlé et mécaniquement stable du fuseau (Microtubule organization by the antagonistic mitotic motors kinesin-5 and kinesin-14 2010).
1. Les kinésines de la famille kinésine-14, principalement HSET (KIFC1) chez les mammifères et Ncd chez la drosophile, jouent un rôle central dans la génération de forces antagonistes centripètes.
En se déplaçant vers l’extrémité (-) des microtubules et en reliant des microtubules antiparallèles, elles s’opposent mécaniquement à l’éloignement des pôles.

(Figure : vetopsy.fr d'après Ogren et coll)
2. En s’opposant aux forces d’expansion produites par Eg5/KIF11 et renforcées par KIF15, les kinésines-14 limitent l’écartement excessif des pôles, préviennent la sur-extension du fuseau et contribuent à la robustesse mécanique globale de la structure bipolaire.
L’action des kinésines-14 ne bloque pas l’anaphase B mais en module l’amplitude et la régularité, en assurant que l’allongement du fuseau reste compatible avec la cohésion de la zone médiane et la transition ultérieure vers la télophase.
Forces externes au fuseau central
Bien que s’exerçant en dehors de la zone médiane, les forces de traction exercées par la dynéine corticale sur les microtubules astraux constituent une composante majeure de l’anaphase B.
1. Elles tirent activement les pôles du fuseau vers le cortex cellulaire, amplifiant l’éloignement des masses chromosomiques.
2. Les deux systèmes de forces ne sont pas redondants.
- Les forces d'expansion internes poussent depuis le centre.
- Les forces de traction externes tirent depuis la périphérie.
La combinaison du raccourcissement des microtubules kinétochoriens et de l’allongement du fuseau polaire au cours de l’anaphase garantit une progression régulière, simultanée et symétrique des chromatides, assurant un partage équitable du matériel génétique et préparant l’entrée en télophase.
Régulations moléculaires de l'anaphase
1. L’activation de l’APC/CCdc20 constitue le signal déclencheur unique de l’anaphase.
a. La levée définitive du point de contrôle du fuseau (SAC) libère Cdc20, permettant l’activation complète de l’APC/C.
- La dégradation rapide de la sécurine libère la séparase, qui clive la cohésine centromérique et initie l’individualisation des chromatides sœurs.
- Ce clivage coordonné impose une entrée synchrone en anaphase pour l’ensemble des chromosomes.
b. Cette activation constitue un point de non-retour, engageant irréversiblement la cellule dans la phase de ségrégation.
2. L’ubiquitination de la cycline B par l’APC/CCdc20 cible le complexe cycline B/CDK1 (MPF) à la dégradation protéasomale.
La diminution progressive de l’activité MPF accompagne la poursuite de l’anaphase et prépare les événements de la télophase, sans intervenir dans le déclenchement initial de la séparation des chromatides.
3. En parallèle de l’activation de l’APC/C, les kinases mitotiques Plk1, Aurora A et Aurora B participent à la coordination spatiale et à la dynamique du fuseau en anaphase ( fuseau en anaphase).
Passage à la télophase
La transition vers la télophase s’effectue lorsque les chromosomes atteignent pleinement les pôles et que débute la reformation de l’enveloppe nucléaire autour de chaque lot chromosomique.
Biologie cellulaire et moléculaireConstituants de la celluleReproduction cellulaireMatériel génétiqueFuseauCycle cellulaireInterphasePhase G1Phase SPhase G2MitoseProphasePrométaphaseMétaphaseAnaphaseTélophaseCytokinèseAbcissionMéioseRégulation du cycle cellulaireCyclinesCDKComplexes Cyclines/CDKPoints de contrôles (checkponts)Enzymes mitotiquesMoteurs mitotiquesComplexes SMC (condensines et cohésines)BiochimieTransport membranaire Moteurs moléculairesVoies de signalisation