La dynamique du fuseau mitotique évolue au cours de la mitose par un remodelage coordonné des microtubules et des forces, permettant l’alignement, la séparation et la réorganisation finale des chromosomes.
La dynamique des microtubules du fuseau varie selon la phase de la mitose, en lien direct avec les fonctions mécaniques attendues à chaque étape.
Elle repose sur une instabilité dynamique marquée des extrémités (+), alternant polymérisation et dépolymérisation, permettant des variations rapides de longueur et une réorganisation continue du réseau microtubulaire.
Cette dynamique est étroitement couplée à la génération et au maintien de la tension bipolaire, qui émerge des propriétés mécaniques combinées des microtubules, de leur organisation spatiale et de leurs interactions avec les chromosomes et les pôles du fuseau.
Instabilité des microtubules selon les phases mitotiques
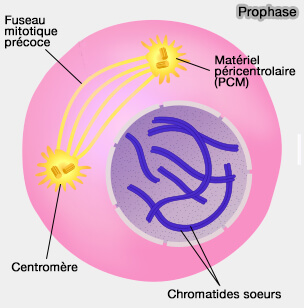
Les centrosomesdupliqués en phase S et maturés en phase G2 migrent vers des pôles opposés, déclenchant l’extension radiale des microtubules et la bipolarisation progressive du fuseau.
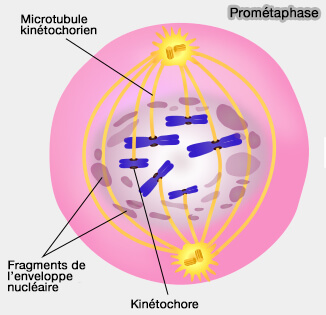
En prométaphase, les chromosomes sont dispersés dans le volume cellulaire et les kinétochores ne sont pas encore engagés de manière stable avec le fuseau.
1. La rupture de l’enveloppe nucléaire rend les kinétochores accessibles aux microtubules, marquant le début de la capture chromosomique.
La forte instabilité dynamique des microtubules du fuseau favorise l’exploration de l’espace cellulaire.
L’allongement des microtubules permet aux extrémités (-) d’atteindre des chromosomes éloignés des pôles et d’entrer en contact avec les kinétochores.
La dépolymérisation rapide empêche la stabilisation prématurée d’attachements incorrects et autorise la répétition de cycles d’exploration.
2. La capture des chromosomes s'effectue en deux temps.
a. Dans un premier temps, les chromosomes sont capturés par des attachements latéraux, au cours desquels les kinétochores interagissent avec la paroi des microtubules.
Ces interactions sont transitoires et dépendent de moteurs associés au fuseau, permettant le repositionnement des chromosomes et leur orientation par rapport aux pôles.
Les kinétochores présentent alors une corona fibreuse développée, structure transitoire qui augmente la surface d’interaction avec les microtubules et favorise les attachements latéraux dynamiques.
b. Dans un deuxième temps, ces attachements latéraux commencent à être convertis en attachements end-on stables, dans lesquels l’extrémité (+) du microtubule s’engage directement dans l’interface kinétochorienne, processus qui se poursuit et se stabilise en métaphase.
Prométaphase
(Figure : vetopsy.fr d'après Ali Zifan)
Cette conversion s’opère sous l’effet de la tension générée par l’engagement bipolaire des chromosomes, favorisant la sélection et la consolidation progressive des attachements corrects, tandis que les attachements erronés sont activement corrigés.
Ce processus permet la maturation fonctionnelle de l’architecture bipolaire du fuseau et l’intégration complète des chromosomes dans son système mécanique.
c. À mesure que les attachements end-on stables s’établissent et que la tension bipolaire augmente, on assiste à une résorption de la corona fibreuse qui accompagne :
la maturation mécanique du fuseau,
la stabilisation des interfaces kinétochore-microtubule,
la transition vers un état métaphasique caractérisé par des attachements verrouillés.
3. À ce stade, les kinétochores non attachés ou incorrectement attachés activent le point de contrôle du fuseau (SAC), assurant la surveillance de l’état d’attachement et empêchant la progression prématurée vers la métaphase tant que la biorientation et une tension bipolaire adéquate ne sont pas établies.
Prométaphase zone médiane courte, dense, non extensible, PRC1 + KIF4A → verrouillage mécanique, forces absorbées → pas d’allongement.
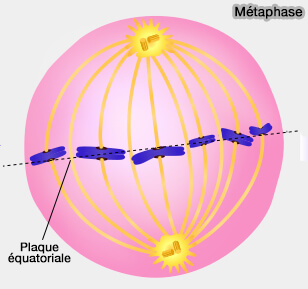
Métaphase
En métaphase, la dynamique des microtubules associés aux chromosomes est partiellement stabilisée, permettant :
la génération d’une tension bipolaire soutenue, tout en conservant une capacité d’ajustement nécessaire à la correction d’erreurs.
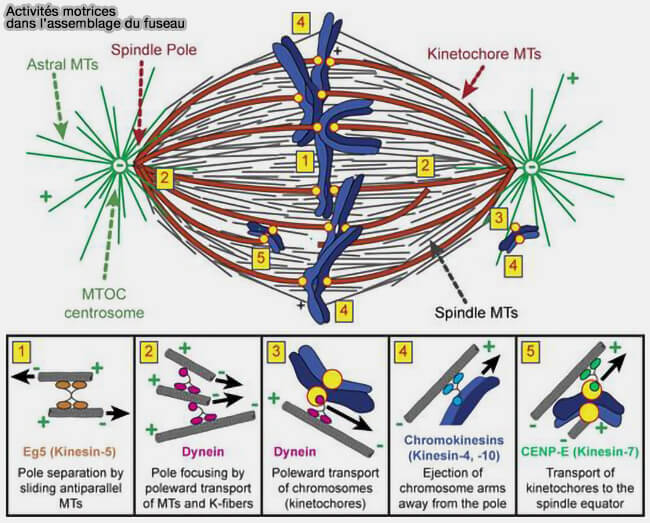
1. À ce stade, les microtubules kinétochoriens et polaires établissent un équilibre de forces opposées qui stabilise l’architecture bipolaire du fuseau et maintient les chromosomes alignés sur la plaque équatoriale.
Les kinésines-14 (HSET/KIFC1) génèrent des forces centripètes qui limitent l’expansion interpolaire excessive du fuseau.
Toutefois, les forces d'expansion sont équilibrées par des forces de traction dépendantes de la dynéine exercées aux pôles du fuseau et sur les microtubules astraux, contribuant au maintien de la distance entre les pôles et à la stabilité globale du fuseau.
Cet équilibre entre forces d’expansion interpolaire et forces de traction polaires stabilise la distance entre les pôles et maintient une géométrie bipolaire compatible avec l’alignement métaphasique des chromosomes.
Remarque : l’architecture et la cohésion du chevauchement antiparallèle impliquent également des régulateurs de la zone médiane, notamment KIF4A, les kinésines-6 MKLP1/KIF23 et MKLP2/KIF20A).
Attachements kinétochoriens
(Figure : vetopsy.fr d'après Heald et Khodjakov)
2. Cette stabilisation locale repose également sur des MAP (protéines associées aux microtubules) qui renforcent la densité et la persistance des microtubules du fuseau en métaphase, en particulier à proximité des chromosomes.
a. TPX2 contribue à cette stabilité en renforçant le réseau microtubulaire périchromosomique, issu de la nucléation locale dépendante de Ran-GTP ( rôles de TPX2).
TPX2 contribue à la nucléation et à la stabilisation des microtubules dans la région chromosomique du fuseau, agissant comme un facteur d’intégration locale sans intervenir directement dans les forces d’écartement des pôles.
En augmentant la persistance des microtubules formés autour des chromosomes et en favorisant la sélection d’orientations compatibles avec les axes du fuseau, TPX2 participe à la robustesse mécanique du réseau central.
Cette action ne rigidifie pas le fuseau, mais soutient une architecture suffisamment dense et cohérente pour maintenir les attachements kinétochoriens end-on sous tension et stabiliser l’alignement métaphasique.
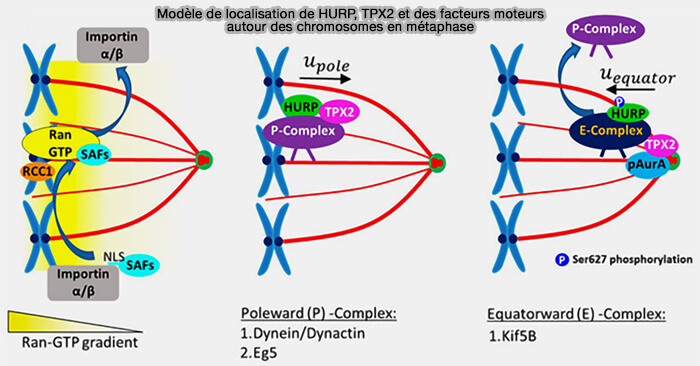
Remarque : un gradient de Ran-GTP libère des facteurs d’assemblage du fuseau (SAFs), tels que HURP, TPX2 et Eg5.
Modèle de localisation de HURP, TPX2 et des facteurs moteurs en métaphase
(Figure : vetopsy.fr d'après Didascalou et col)
a. En augmentant la persistance des fibres périchromosomiques et en limitant les catastrophes locales, HURP consolide les microtubules impliqués dans la capture, l’alignement et le maintien des chromosomes sur la plaque équatoriale.
Cette stabilisation régionale améliore l’efficacité des interactions kinétochore-microtubule et soutient la correction et la maintenance des attachements end-on sous tension, tout en préservant une dynamique suffisante pour l’ajustement fin des fibres.
b. Cette stabilisation locale est renforcée par la coordination de HURP avec la kinésine-8 KIF18A, qui contribue à amortir la dynamique des extrémités (+) des microtubules kinétochoriens et à maintenir un alignement métaphasique précis.
Alors que TPX2 agit principalement en amont, en organisant et en densifiant le réseau microtubulaire périchromosomique naissant, HURP intervient plus localement en aval pour stabiliser et ajuster la dynamique des microtubules déjà engagés dans l’alignement métaphasique.
Remarque : bien que TPX2 et HURP agissent toutes deux à proximité des chromosomes, TPX2 relève des MAP
organisatrices
qui structurent le réseau microtubulaire périchromosomique, tandis que HURP appartient aux MAP
stabilisatrices qui modulent finement la dynamique des fibres déjà établies.




{kind=link}
 rôles de TPX2).
rôles de TPX2).