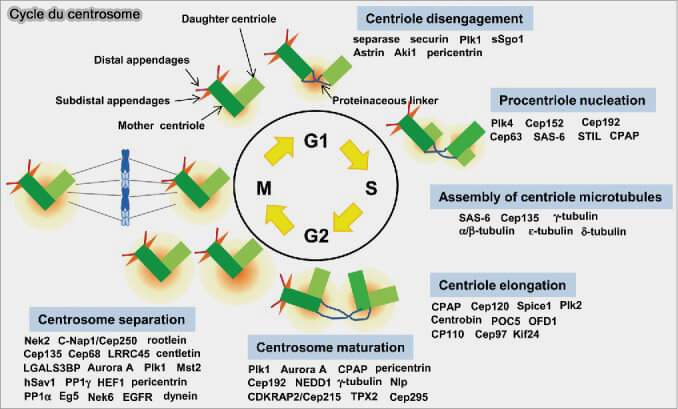
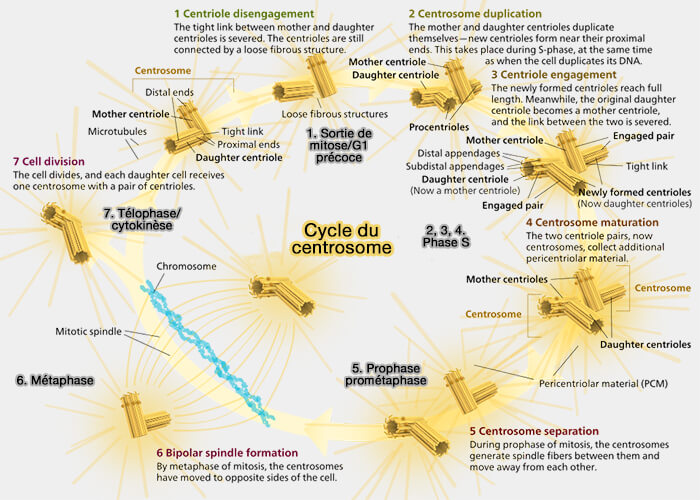
Le cycle du centrosome intègre la duplication contrôlée des centrioles, l’expansion du PCM et la séparation coordonnée des pôles pour assurer la formation d’un fuseau bipolaire fiable.
Le cycle du centrosome repose sur l’enchaînement coordonné de plusieurs processus :
Le désengagement crée la possibilité structurelle de dupliquer, mais n’autorise pas la duplication.
Remarque : dans les schémas simplifiés, ce processus est parfois représenté par des " fibres de connexion ", mais il s’agit d’une convention graphique : aucune fibre réelle n’unit les deux centrioles après la mitose, i.e. la proximité est uniquement due au PCM partagé.
2. Ce processus dépend de l’activité combinée de Plk1, de la séparase et du remodelage des composants proximaux tels que SAS-6 et STIL.
Plk1 phosphoryle plusieurs éléments du lien proximal et affaiblit l’architecture du cartwheel.
La séparase clive des substrats encore mal définis impliqués dans la cohésion mère-fille.
En parallèle, la dégradation ou la dissociation progressive de SAS-6 et STIL démantèle le cartwheel, ce qui supprime définitivement l’interface qui retenait le centriole fille sur la face proximale du centriole mère.
Fin de G1 : licence en et préparation du centriole mère
La licence (centriole licensing) est l’ensemble des conditions moléculaires qui autorisent un centriole à initier une nouvelle duplication au cours de la phase S.
1. Chaque centriole mère génère un procentriole, ce qui aboutit à deux paires mère-fille, mais le centrosome reste unique, car les deux paires partagent un seul PCM commun ( duplication du centriole).
Le procentriole se forme orthogonalement au centriole mère et demeure engagé à celui-ci tout au long du cycle, conformément au maintien du lien proximal.
2. Parallèlement, le PCM commence à se réorganiser :
CEP295, bien que surtout impliqué dans la conversion fille/mère en G1 du cycle suivant, commence effectivement à s’accumuler dès la phase S.
augmentation progressive de sa capacité de recrutement des complexes γ-TuRC, conditionnant la future compétence nucléatrice du centrosome.
3. Le relais du complexe cycline A/CDK2 soutient l’allongement des procentrioles et la densification initiale du PCM, assurant la progression harmonieuse du programme structural de duplication.
Phase G2 : maturation centrosomale
Au cours de la phase G2, les centrosomes subissent une maturation progressive, principalement portée par l’expansion et la réorganisation du matériel péricentriolaire (PCM), qui accroît fortement leur capacité à nucléer les microtubules en vue de la mitose.
1. Durant la phase G2, sous l’effet des kinases mitotiques, le PCM subit une expansion radiale marquée destinée à :
accroître la surface active du centrosome,
initier rapidement la formation des microtubules du fuseau,
CEP192 joue ici un rôle pivot dans l’organisation de la plateforme γ-TuRC et dans l’assemblage de la matrice du PCM, tandis que CEP152, bien que principalement impliqué dans l’ancrage et la régulation de PLK4 en G1/S, contribue indirectement à un PCM correctement structuré en stabilisant l’architecture proximale du centriole mère sur laquelle le PCM se déploie.
l’arrivée de modules coiled-coil supplémentaires pour renforcer la charpente, issus de composants structuraux tels que PCNT, CDK5RAP2 (CEP215) et CEP295 ou rootletin (CROCC), qui s’assemblent en un réseau fibrillaire dense renforçant la charpente mécanique du PCM et stabilisant son expansion radiale,
le recrutement massif des γ-TuRC, dont la multiplication des sites d’ancrage confère au centrosome une capacité nucléatrice maximale en vue de l’entrée en mitose.
2. En parallèle, le centrosome devient un hub d’activation des régulateurs G2/M, i.e. principalement Plk1, Aurora A et, en fin de G2, la montée progressive du complexe cycline B/CDK1 (MPF), dont l’activation coordonnée conditionne la transition vers la prophase et l’expansion finale du PCM ( plateforme des kinases).
3. À la fin de la G2, les deux ensembles mère-fille possèdent chacun un PCM densifié et fonctionnellement équivalent, mais demeurent encore appariés en une seule unité.
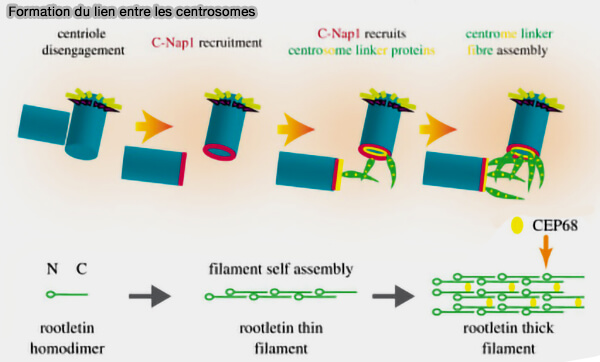
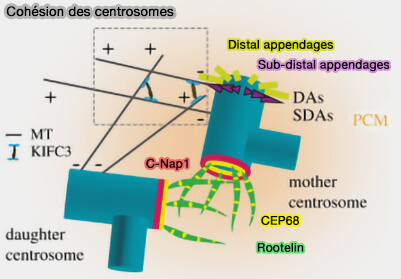
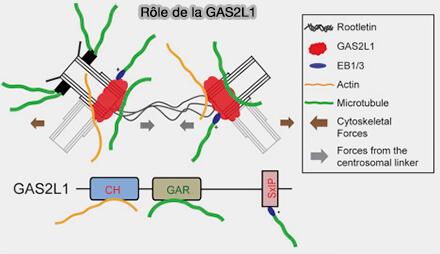
Cohésion des centrosomes
(Figure : vetopsy.fr d'après Dang et Schiebel) CEP250 (C-Nap1) fixé à la base des centrioles mères, et constituant l’ancrage proximal du lien,
GAS2L1, qui interagit avec C-Nap1 et contribue à la stabilité du lien intercentriolaire en reliant la cohésion proximale des centrioles aux réseaux d’actine et de microtubules.
Remarque : la dynéine est le moteur majeur des forces de tension et de cohésion entre les centrosomes ( cf. figure-ci-dessous).
Toutefois, KIFC3, kinésine à polarité (-), se déplaçant à l’inverse des kinésines classiques, peut générer des forces de glissement entre microtubules interphasiques, tandis que la dynéine constitue le moteur principal assurant les forces de cohésion et de positionnement des centrosomes.
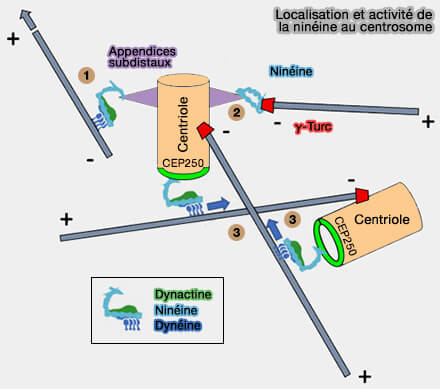
Localisation et activité de la ninéine au centrosome
(Figure : vetopsy.fr d'après Guo et Merdes)
b. Le maintien de l’appariement garantit un unique réseau microtubulaire radial, assurant :
un MTOC (centre organisateur des microtubules) unique,
En entrée de mitose, i.e. G2 ➞ M, les deux unités centrosomales (chaque unité = centriole mère + centriole fille) doivent se séparer pour former les pôles du fuseau bipolaire.
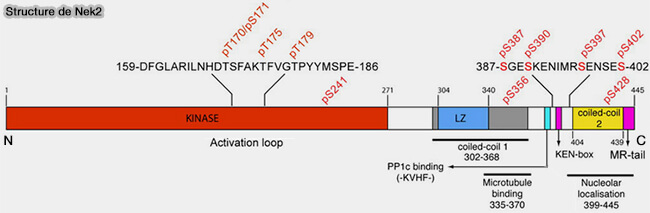
Structure de Nek2
(Figure : vetopsy.fr d'après Rellos et coll)
affaiblissant l’ancrage du linker proximal aux réseaux d’actine et de microtubules.
Remarque : durant la prophase, Plk1 amplifie le processus en phosphorylant plusieurs composants du PCM et en facilitant la séparation des deux futurs pôles.
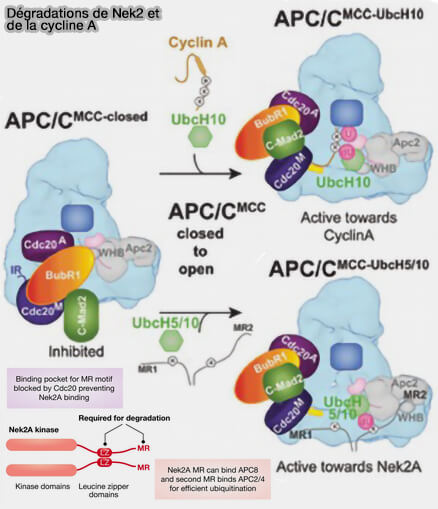
La cycline A est dégradée en premier car sa D-box est accessible dès la prométaphase, puis Nek2A suit immédiatement, ses motifs canoniques de reconnaissance par l’APC/C, i.e. D-box et KEN-box, étant exposés et non protégés.
Dégradation de Nek2 et de la cycline A
(Figure : vetopsy.fr d'après Alfieri et coll)
Cette dégradation précoce de Nek2A est en outre facilitée par un mode de reconnaissance non canonique reposant sur un motif MR, i.e. méthionine (M) et arginine (R), C-terminal, associé au MCC (Mitotic Checkpoint Complex), expliquant ainsi pourquoi sa dégradation peut débuter alors que le SAC (point de contrôle du fuseau mitotique) n’est pas encore complètement levé.
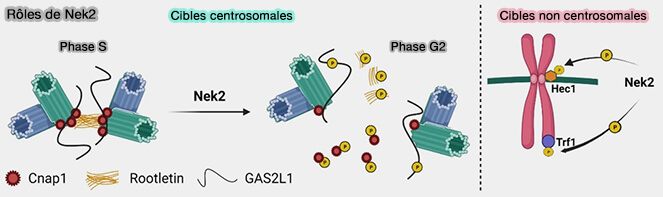
a. La dégradation de Nek2A inhibe sa fonction de phosphorylation sur les cibles centrosomales en :
verrouillant la séparation centrosomale déjà initiée, une fois le lien centrosomal dissous,
empêchant le reclustering des centrosomes, i.e. la reformation anormale d’un pôle unique à partir de plusieurs centrosomes sous l’effet de forces microtubulaires instables, phénomène fréquent dans les cellules cancéreuses multipolaires,
stabilisant la formation du fuseau bipolaire, en maintenant durablement deux pôles distincts et fonctionnels.
L’inactivation, la dissociation ou la dégradation coordonnée de protéines telles que HURP, TPX2 ou KIF18A participe à la perte de la stabilisation métaphasique et à la mise en place des dynamiques propres à l’anaphase.
Ce processus est indépendant du lien mère-fille, lequel reste intact ( lien mère-fille).
Cette modulation des forces exercées le long des microtubules et sur les chromosomes contribue à prévenir le regroupement aberrant des pôles et à stabiliser la bipolarité du fuseau.
Principaux régulateurs de G2/M
Les régulateurs de la transition G2/M coordonnent la préparation mécanique du centrosome en vue de la séparation des pôles et de la formation du fuseau bipolaire.
Plk1 (Polo-like kinase 1) amplifie la séparation des centrosomes initiée en fin de G2, en coordonnant l’expansion du PCM, l’activation des forces d’écartement et la formation d’asters mitotiques robustes.
Aurora A, activée au centrosome, stabilise l’organisation des asters et facilite leur interaction avec le cortex, contribuant à l’ancrage et au positionnement correct des pôles mitotiques.
Le complexe cycline B/CDK1 (MPF) déclenche la réorganisation globale du PCM en fin de G2, en augmentant sa densité structurale et sa capacité de nucléation microtubulaire, condition préalable à la formation d’asters fonctionnels.
Destins des centrosomes après la séparation
Après leur disjonction en entrée de mitose, les deux centrosomes s’individualisent et deviennent les pôles en formation du futur fuseau bipolaire.
Leur migration, leur activité nucléatrice et leur rôle structural évoluent de manière coordonnée tout au long de la prophase, de la prométaphase et de la métaphase, jusqu’à la ségrégation en anaphase puis la cytokinèse.
1. En prophase, la séparation des deux centrosomes est ensuite amplifiée par la dynamique microtubulaire afin de permettre leur migration active vers deux futurs pôles mitotiques.
La dynéine corticale et les forces d’écartement tirent les centrosomes vers des positions opposées.
Le réseau d’actine contribue à la mise en tension du cytosquelette.
2. En métaphase, chaque centrosome stabilise un pôle du fuseau et maintient la géométrie bipolaire nécessaire à l’alignement des chromosomes.
Le PCM reste densifié, soutenant une production continue de microtubules.
Les asters assurent l’ancrage cortical et contribuent à la mise en tension du fuseau.
Les microtubules astraux et interpolaires organisent un réseau mécanique permettant d’équilibrer les forces opposées exercées sur les chromosomes.
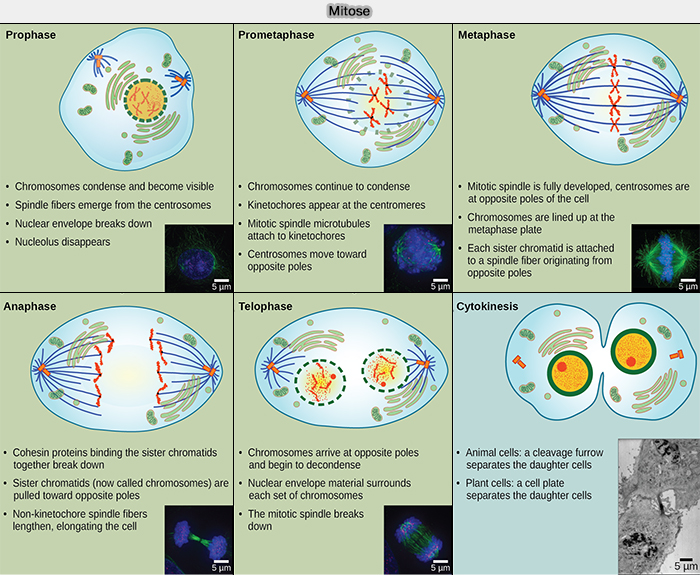
Les phases de la mitose
(Figure : vetopsy.fr d'après Opened-cuny.edu)
3. En anaphase, les centrosomes deviennent des plateformes mécaniques qui guident l’allongement axial du fuseau et participent activement à la ségrégation chromosomique.
Ils s’éloignent l’un de l’autre tout en émettant des microtubules astraux dirigés vers le cortex.
Les microtubules astraux assurent la traction corticale et contribuent à séparer les pôles en amplifiant la distance.
4. En télophase, les centrosomes commencent à perdre leur activité mitotique et préparent la cellule à la transition vers la cytokinèse et l’interphase suivante.
La densité du PCM diminue progressivement.
La capacité nucléatrice décline, réduisant la formation de nouveaux microtubules.
Les asters se rétractent, permettant la réorganisation des réseaux cytosquelettiques autour des deux futurs noyaux.
5. Pendant la cytokinèse, les deux centrosomes jouent un rôle de guidage spatial, mais ne dirigent plus la dynamique du fuseau.
Ils se repositionnent symétriquement dans chaque cellule fille en formation.
Leur activité nucléatrice devient minimale et le réseau astral régresse.
Ils coordonnent indirectement le placement du plan de clivage en interagissant avec les réseaux d’actine périphériques et la contraction de l’anneau de cytokinèse.
Chaque centrosome se prépare à redevenir le MTOC (centre organisateur des microtubules) principal de la cellule en interphase.
5. Au terme de la division, chaque cellule fille hérite d’un centrosome complet contenant :
un centriole mère mature portant des appendices,
un centriole fille issu de la duplication précédente.
Le PCM retrouve une organisation plus compacte et moins dense qu’en mitose, recentré à proximité du noyau.
Le centrosome assume alors à nouveau sa fonction de MTOC unique, organisant le réseau radial interphasique.
Défaillances de la duplication et de la séparation des centrosomes
Un défaut d’appariement ou une disjonction incorrecte perturbe directement l’établissement du fuseau et conduit à des erreurs de ségrégation.
Une disjonction prématurée produit des fuseaux multipolaires, favorisant les aneuploïdies et les instabilités génomiques.
Une disjonction retardée conduit à des fuseaux monopôles, entraînant un arrêt prolongé du cycle via le SAC.
Des défauts de CEP250, rootletin ou Nek2 provoquent une amplification anormale des centrosomes et des divisions défectueuses.





{kind=link}

 lien mère-fille).
lien mère-fille).