Le centriole est un organite cylindrique organisé en neuf triplets de microtubules dont l’architecture proximale, latérale et distale assure la stabilité du cylindre et conditionne ses fonctions cellulaires.
Le centriole est un organite cylindrique hautement spécialisé, organisé en neuf triplets de microtubules et doté d’une architecture tridimensionnelle asymétrique qui conditionne sa fonction.
dirige la nucléation d’un réseau microtubulaire organisé,
constitue le gabarit indispensable à la formation du cil primaire.
Sa configuration modulaire, répartie entre un pôle proximal, une paroi latérale et un pôle distal, assure à la fois stabilité mécanique et spécificité fonctionnelle.
Structure du centriole
Vue d'ensemble
1. La structure fondamentale du centriole repose sur un cylindre d’environ 450-500 nm de long et 200-250 nm de diamètre, formé de neuf triplets de microtubules (A, B et C) disposés en cercle.
a. Chaque triplet comprend :
un microtubule complet (A), formé de 13 protofilaments.
deux microtubules incomplets (B et C) de 10 protofilaments partageant des parois, formant une structure stable capable de servir de base mécanique au centrosome.
b. Les triplets centriolaires, bien que composés d’hétérodimères α/β-tubuline comme les microtubules, sont des structures non dynamiques.
2. La cartographie moléculaire du centriole est en constante évolution :
Les avancées récentes en microscopie haute résolution et en protéomique identifient régulièrement de nouveaux composants dans les modules proximaux, latéraux et distaux, enrichissant et révisant progressivement le modèle canonique présenté ici.
Complexité de l’architecture du centriole selon l’état des connaissances en 2020
(Figure : vetopsy.fr d'après Laporte et coll)
Extrémité proximale
L’extrémité proximale, correspondant à l’extrémité (-) des microtubules du triplet, constitue l’origine architecturale du centriole et le site d’assemblage du procentriole. qui regroupe plusieurs modules structuraux solidaires.
CEP est l’abréviation de CEntrosmal Protéin, tandis que le nombre associé indique la masse moléculaire approximative de la protéine en kDa.
verrouiller la base du centriole, en ancrant fermement les triplets au moyeu SAS-6 via la pinhead et CEP135,
maintenir une géométrie fixe, imposée par le cartwheel et transmise aux triplets par la pinhead,
empêcher toute dynamique d’allongement, grâce à la rigidification assurée par la couronne proximale et l’échafaudage luminal.
Remarque : cette architecture proximale est ensuite fortifiée par des renforts latéraux spécialisés, i.e. le A-C linker, le D2-rod et le module CEP120/SPICE1 ( paroi latérale du centriole).
Structure de l'extrémité proximale du centriole
(Figure : vetopsy.fr d'après LeGuennec et coll)
Cet échafaudage s’adosse étroitement à la face interne des triplets microtubulaires, renforçant la paroi interne et stabilisant la courbure des protofilaments, ce qui prévient leur désorganisation lors de l’allongement et de la maturation du centriole.
Modèle de l’organisation des protéines de l’échafaudage interne du centriole
(Figure : vetopsy.fr d'après Sala et coll)
Elle est distribuée sur l’ensemble de la longueur du centriole, avec un enrichissement proximal, et constitue un élément central du scaffold interne dès les premières étapes de l’assemblage.
POC1 participe au maintien de l’intégrité longitudinale et sert de plateforme structurale pour l’organisation d’autres composants intraluminaux.
2. POC5 est un facteur de maturation tardive, recruté après la formation initiale du procentriole.
Son accumulation est progressive en G2 puis en mitose, en parallèle de l’allongement axial du centriole.
POC5 se localise préférentiellement dans la région intermédiaire à distale de la lumière centriolaire, où elle contribue au renforcement du scaffold interne et à la stabilisation longitudinale du cylindre en cours de maturation.
WDR90/POC16
(Figure : vetopsy.fr d'après Steib et coll)
Elle se localise le long de la paroi interne des triplets microtubulaires, au contact direct du mur microtubulaire, avec un enrichissement marqué dans la région distale de la lumière centriolaire.
WDR90 participe au maintien de l’intégrité architecturale du centriole et s’intègre au réseau intraluminal longitudinal formé par POC1 et POC5, contribuant à la rigidité et à la stabilité mécanique du cylindre.
4. Le rôle porteur majeur du scaffold intraluminal repose principalement sur l’axe POC1-POC5-WDR90, mais d'autres protéines internes intraluminales, visibles sur les figures, contribuent à l’organisation spatiale, à la maturation et à la stabilité globale du cylindre centriolaire.
La centrine (CETN), protéine intraluminale servant de repère axial et de maturation, est localisée dans la lumière centriolaire sans rôle porteur majeur dans le scaffold interne.
MDM1, composant intraluminal associé à la paroi interne des triplets, contribue à l’organisation disto-intermédiaire et à la maturation du centriole.
FAM161A, protéine du réseau intraluminal longitudinal, participe à la stabilité globale et à la cohésion de l’architecture interne.
WD40A et WD40B, protéines à domaines WD40 distinctes de WDR90, sont associées à la lumière centriolaire et impliquées dans l’organisation de complexes intraluminaux sans constituer l’armature principale.
Extrémité distale
L’extrémité distale, correspondant à l’extrémité (+) des microtubules du triplet, forme une coiffe protectrice composée de plusieurs protéines qui bloquent totalement l’accès à la β-tubuline terminale.
Structure de l'extrémité distale du centriole
(Figure : vetopsy.fr d'après LeGuennec et coll)
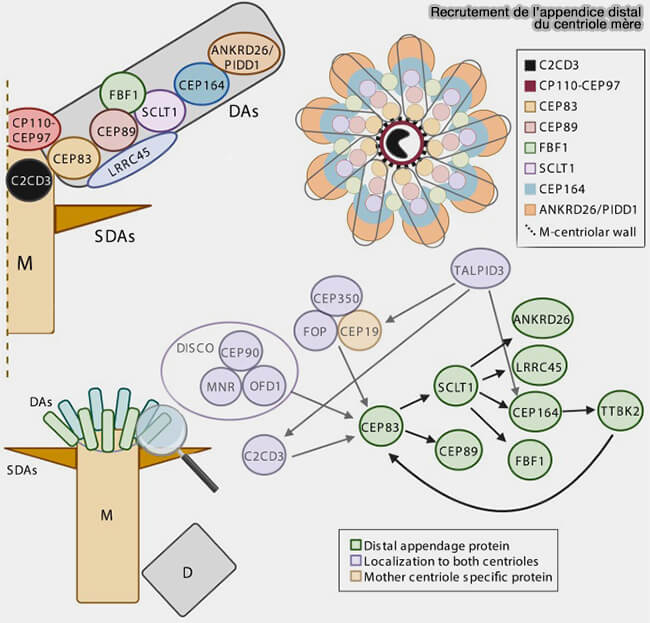
1. Les appendices distaux (distal appendages ou DA), i.e. CEP164, CEP83, CEP89, SCLT1 et FBF1, s’attachent à l’extérieur du tubule A et verrouillent la fin des triplets, définissant le site d’ancrage du cil primaire.
Ces appendices, formés par ODF2, la ninéine et CEP170, constituent la plateforme structurale sur laquelle s’organisent les mécanismes d’ancrage des microtubules cytoplasmiques.
Appendices subdistaux du centriole mère (ciliogenèse)
(Figure : vetopsy.fr d'après Huang et coll)
3. La coiffe terminale (capping complex/distal cap) est composée de plusieurs protéines.
ODF2 contribue à la rigidification du socle distal.
La zone de transition (transition zone), i.e. TMEM67, TMEM216, NPHP1, 3, 4 (néphrocystines), MKS1, forme un anneau qui scelle encore davantage l’extrémité distale.
4. Une densité annulaire appelée " torus ", visible en cryotomographie autour du cartwheel, renforce la base des triplets en stabilisant la jonction spoke-pinhead-tubule A.
Sa composition moléculaire exacte reste toutefois indéterminée, cette structure correspondant pour l’heure à une signature morphologique plutôt qu’à un module protéique identifié et pourrait être associée à :
la coiffe terminale,
des composants proximaux des appendices distaux,
possiblement CEP162/CEP290 selon les espèces et stades.
5. La fonction de l'extrémité distale est de :
fermer hermétiquement l’extrémité distale,
empêcher toute dynamique terminale, du fait de l’absence de coiffe GTP active qui rend impossible tout allongement, contrairement aux microtubules cytoplasmiques ( dynamique des microtubules).
préparer l’interface avec la ciliogenèse.
Paroi latérale
La portion latérale des triplets, i.e. jonction A/B/C, est stabilisée par des protéines dédiées, essentielles pour maintenir l’intégrité mécanique du cylindre.
relie le tubule A d’un triplet au tubule C du triplet voisin,
assure la cohésion radiale du cylindre,
stabilise l’organisation x 9.
Lien A-C du centriole
(Figure : vetopsy.fr d'après Cai et coll)
2. La tige D2 (D2-rod) longitudinale de renfort observée en cryo-ET, qui s’étend le long de la paroi externe des triplets, participe au maintien de la rigidité longitudinale du cylindre.
4. D'autres stabilisateurs latéraux centriole scaffold comme CEP295 et POC1/POC5 complètent l’armature interne pour maintenir la continuité radiale.
4. La fonction de la paroi latérale est de :
consolider la jonction A-B-C,
empêcher l’ouverture des protofilaments,
garantir une cohésion radiale stable grâce au lien A-C,
maintenir les triplets dans un état mécaniquement figé.
L’ensemble de ces modules, i.e. cartwheel proximal, stabilisation latérale par CEP120/SPICE1 et coiffe distale CP110/CEP97/CEP164, ferme les extrémités et verrouille les parois du triplet, rendant les microtubules centriolaires non dynamiques et incapables d’allongement.









 paroi latérale du centriole).
paroi latérale du centriole).