Le centrosome assure l’ancrage et la stabilisation des microtubules cytoplasmiques grâce aux appendices subdistaux du centriole mère, en convertissant la nucléation en un réseau radial mécaniquement cohérent.
Les appendices subdistaux sont fixés à la paroi externe du tubule A, immédiatement sous les appendices distaux.
Appendices subdistaux du centriole mère (ciliogenèse)
(Figure : vetopsy.fr d'après Huang et coll)
1. Les appendices subdistaux sont des structures protéiques constitutives du centriole mère, dérivées de la paroi externe du tubule A et intégrées au matériel péricentriolaire (PCM), formées principalement par trois protéines.
b. La ninéine, localisée plus distalement et impliquée dans l’interface avec le réseau microtubulaire, constitue l’élément clé de l’interface fonctionnelle avec les microtubules cytoplasmiques.
Il participe à la charpente coiled-coil de l’appendice subdistal et en détermine la longueur, l’orientation et la géométrie.
Il coopère avec la ninéine pour organiser l’extrémité fonctionnelle de l’appendice, en stabilisant la plateforme sur laquelle la ninéine exerce l’ancrage des extrémités (-) des microtubules.
Il est requis pour une organisation radiale correcte des microtubules ancrés au centrosome, i.e. en son absence, les microtubules peuvent être nucléés et partiellement ancrés, mais le réseau est mal orienté et désorganisé.
2. Ces appendices constituent la plateforme structurale coiled-coil stable, visible en microscopie électronique, sur laquelle s’organisent les mécanismes d’ancrage des microtubules cytoplasmiques :
sans interagir avec les triplets centriolaires eux-mêmes,
indépendamment de la présence de microtubules ancrés.
3. Les appendices subdistaux recrutent des adaptateurs moteurs qui convertissent la plateforme structurale en site fonctionnel d’ancrage et de traction des microtubules (Who are you, subdistal appendages of centriole? 2018).
SDA (Spindle Defective Assembly) est qualifiée d’adaptateur moteur car elle relie la plateforme structurale des appendices subdistaux à la machinerie motrice dépendante de la dynéine, sans qu'elle constitue elle-même un composant du moteur.
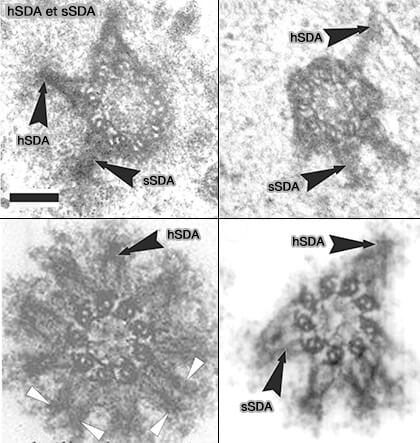
a. hSDA (forme longue de SDA) est localisée aux appendices subdistaux du centriole mère, où elle agit comme adaptateur de la dynéine, favorisant l’ancrage des microtubules à leur extrémité (-) et la génération de forces de traction nécessaires à la stabilisation des asters et à la séparation des centrosomes.
b. sSDA (forme courte), issue d’un épissage alternatif, est une isoforme tronquée qui conserve la localisation aux appendices subdistaux, mais perd les régions C-terminales nécessaires à l’assemblage d’un module moteur complet, notamment via la ninéine.
Il agit comme un modulateur négatif de hSDA en entrant en compétition directe avec hSDA pour les sites d’ancrage disponibles.
sSDA ne permet pas le recrutement stable ou correct de l’ensemble des partenaires moteurs, conduisant à la formation de complexes d’ancrage incomplets.
Les microtubules restent ancrés, mais les forces de traction sont atténuées, leur persistance temporelle réduite et les événements de glissement ou de traction excessive limités.
sSDA agit comme un frein mécanique fin en empêchant une activation prématurée ou excessive des forces dépendantes des appendices subdistaux et en ajustant le seuil à partir duquel la séparation des centrosomes devient possible.
Ninéine et complexe d'ancrage des microtubules
Pour résumer l'ancrage des microtubules au centrosome, la ninéine recrute, Ndel1 connecte, Lis1 stabilise, la dynéine s’attache au microtubule.
Vue d'ensemble
1. Le complexe d’ancrage désigne l’assemblage fonctionnel qui s’organise sur les appendices subdistaux du centriole mère afin d’assurer l’ancrage et l’organisation du réseau microtubulaire cytoplasmique.
L’ensemble ninéine-dynéine assure l’ancrage des microtubules sur la plateforme structurale fournie par les appendices subdistaux, indépendamment de la nucléation par les complexes γ-TuRC, qui relève exclusivement du matériel péricentriolaire (PCM).
Remarque : des MAP associées, telles que MAP7 (Ensconsin), peuvent interagir transitoirement avec ce module pour stabiliser le réseau microtubulaire ancré, sans faire partie du complexe d’ancrage structural proprement dit ( MAP associées au centrosome).
2. Par l’action coordonnée de ces modules :
le réseau microtubulaire maintient sa cohésion autour du centrosome et résiste aux contraintes mécaniques en empêchant la dispersion des microtubules, un rôle confirmé par la désorganisation globale observée lors de la déplétion de la ninéine,
les microtubules adoptent une organisation radiale stable en interphase,
le positionnement du centrosome et l’orientation du fuseau sont correctement préparés avant l’entrée en mitose.
1. La ninéine, localisée à l’extrémité distale des appendices subdistaux, organise l’interface fonctionnelle entre le centriole mère et le réseau microtubulaire cytoplasmique.
1. Elle définit une zone d’ancrage privilégiée où les extrémités (-) des microtubules sont maintenues au voisinage immédiat du centrosome.
Cette zone ne correspond pas à un point de fixation ponctuel, mais à un volume fonctionnel restreint, dans lequel les microtubules sont maintenus proches du centrosome tout en conservant leur intégrité structurale.
a. Au niveau moléculaire, cette organisation n’implique pas la reconnaissance de la tubuline terminale ni l’obstruction de l’extrémité (-), i.e. la ninéine ne possède pas de domaine de liaison spécifique de l’extrémité microtubulaire et n’interagit pas avec les protofilaments terminaux.
À la place, la ninéine favorise la capture du lattice microtubulaire proximal de manière indirecte, en recrutant et en positionnant la dynéine cytoplasmique et ses régulateurs Nde1 et Lis1 (rôle de la dynéine).
Ces composants interagissent directement avec les surfaces latérales de la tubuline polymérisée, situées à proximité de l’extrémité (-), assurant un ancrage mécanique stable sans contact avec la tubuline terminale.
b. Cette capture latérale confère simultanément plusieurs avantages fonctionnels.
Elle préserve la dynamique intrinsèque de l’extrémité (-) en évitant toute perturbation de l’équilibre local de polymérisation et de dépolymérisation des protofilaments.
Elle répartit les contraintes mécaniques sur une surface d’interaction étendue, mécaniquement plus robuste qu’un point d’attache terminal unique.
Elle permet enfin l’application de forces de traction soutenues sans induire de dépolymérisation locale.
2. En organisant spatialement cette interaction latérale, la ninéine impose une géométrie contrainte dans laquelle le microtubule est orienté radialement vers le centrosome et est maintenu à une distance compatible avec l’action de la dynéine.
Rôle de la ninéine
(Figure modifiée : vetopsy.fr d'après Delgehyr et coll)
Cette fonction relève exclusivement du γ-TuRC recruté au PCM par CEP192, NEDD1 et CDK5RAP2.
Elle agit en aval, en stabilisant et en ancrant les microtubules déjà formés aux appendices subdistaux, contribuant ainsi à la cohésion et à l’architecture radiale du réseau microtubulaire.
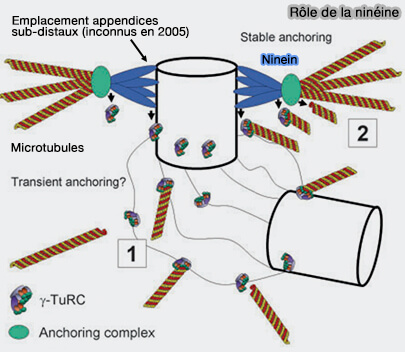
Remarque : dans cette figure de 2005, la localisation de la ninéine indique la zone où se trouvent les appendices subdistaux, dont la structure n’avait pas encore été décrite à l'époque.
Elle ancre et stabilise les microtubules au centriole mère.
Lors de la mitose, elle ne s’associe pas aux microtubules du fuseau, mais son action préalable sur la cohésion radiale et le positionnement du centrosome conditionnent l’orientation correcte du fuseau.
Rôle de la dynéine dans l’ancrage des extrémités (-)
L’ancrage des extrémités (-) des microtubules aux appendices subdistaux du centriole mère ne repose pas sur une interaction directe entre la ninéine et la tubuline, mais sur un mécanisme actif de capture et de mise sous tension, dépendant de la dynéine cytoplasmique et de ses régulateurs.
Principe général de l’ancrage des extrémités (-)
1. Contrairement à l’extrémité (+), qui peut être coiffée par des complexes terminaux, comme les protéines EB et les complexes +TIPs associés, l’extrémité (-) des microtubules interphasiques est stabilisée par un ancrage fonctionnel, impliquant :
une capture latérale du microtubule à proximité de son extrémité (-),
un maintien spatial contre les appendices subdistaux,
une mise sous tension qui empêche la diffusion ou le détachement du microtubule.
Régulation de la dynéine par Ndel1/Nde1
(Figure : vetopsy.fr d'après Garrott et coll)
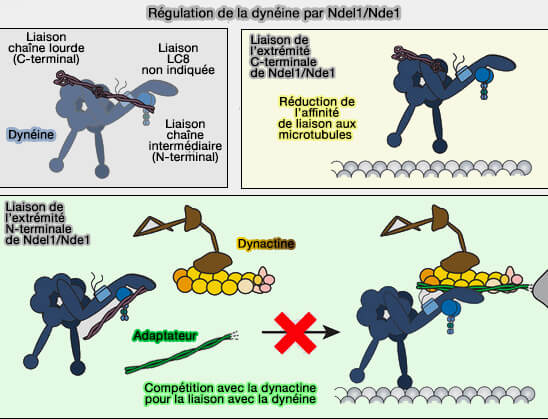
1. Nde1/Ndel1 est un adaptateur connu de la dynéine, capable de se lier :
à des protéines d'assemblage riches en coiled-coil, comme ceux de la ninéine, sans qu'aucune interaction structurale directe ninéine-Ndel1 médiée par un appariement coiled-coil spécifique n’ait été formellement démontrée,
Le recrutement de Ndel1 repose sur sa capacité à relier fonctionnellement la dynéine associée aux microtubules au module d’ancrage des appendices subdistaux, assurant ainsi la continuité mécanique entre le réseau microtubulaire cytoplasmique et la plateforme centriolaire.
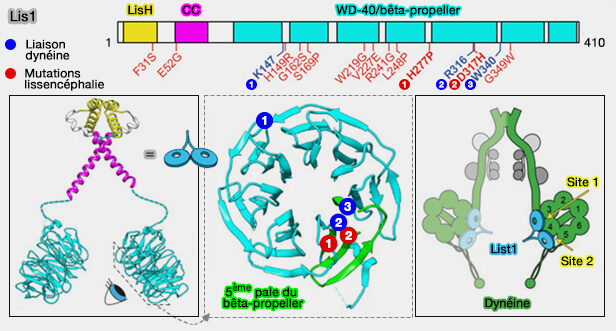
2. Lis1 n’est pas un adaptateur de la ninéine, mais un régulateur de la dinéine, interagissant également avec Ndel1.
Lis1 se lie au domaine moteur AAA+ de la dynéine, la stabilise dans un état fortement associé au microtubule et empêche le décrochage du moteur sous contrainte mécanique.
Lis1 agit donc en aval du recrutement, en modulant la cinétique et la force du moteur, mais pas son adressage.
Structure et liaisons de Lis1
(Figure : vetopsy.fr d'après Markus et coll)
3. Sur le plan mécanistique, cette organisation a plusieurs conséquences immédiates.
Elle induit un enrichissement local de dynéine au niveau des appendices subdistaux, augmentant fortement la densité effective de moteurs disponibles à proximité des extrémités (-).
Elle accroît ainsi la probabilité de rencontre entre la dynéine et le lattice microtubulaire proximal, condition limitante pour l’établissement d’un ancrage stable.
Enfin, elle favorise la stabilisation de complexes dynéine-microtubule à proximité de l’extrémité (-), en maintenant le moteur dans une configuration compatible avec une traction prolongée.
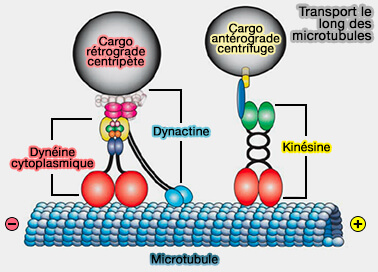
4. La dynéine cytoplasmique, DYNC de type 1, assure la liaison directe au microtubule par son domaine moteur.
Cette interaction s’effectue avec le lattice microtubulaire, indépendamment de l’extrémité terminale, et constitue le point de contact effectif permettant la transmission des forces de traction vers le centrosome.
Remarque : la dynactine n’est pas requise pour la reconnaissance du microtubule elle-même, mais elle module l’organisation, la stabilité et l’efficacité fonctionnelle du complexe moteur lorsqu’une traction soutenue ou une coordination spatiale accrue est nécessaire, par exemple lors du maintien prolongé des microtubules astraux sous tension ou de la résistance aux forces cytoplasmiques opposées.
Pour résumer l'ancrage des microtubules au centrosome, la ninéine recrute, Ndel1 connecte, Lis1 stabilise, la dynéine s’attache au microtubule.
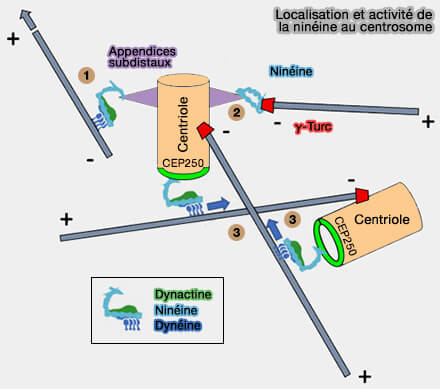
1. La nucléation par le complexe γ-TuRC positionne d’emblée l'extrémité (-) des microtubules au voisinage des appendices subdistaux, condition nécessaire à leur ancrage mécanique ultérieur, tandis que l’extrémité (+) s’allonge vers le cytoplasme.
Les principaux recruteurs du γ-TuRC, i.e. CEP192, NEDD1 et CDK5RAP2, sont ancrés au matériel péricentriolaire (PCM) proximal du centriole mère, ce qui concentre les complexes γ-TuRC au voisinage immédiat des appendices subdistaux, sans que ceux-ci n’interviennent directement dans la nucléation.
Localisation et activité de la ninéine au centrosome
(Figure : vetopsy.fr d'après Guo et Merdes)
2. Dans cette zone, la dynéine-1, recrutée et orientée fonctionnellement par la géométrie imposée par la ninéine et l’appendice subdistal, se lie directement au lattice microtubulaire via son domaine moteur, à proximité de l’extrémité (-).
a. La dynéine-1 hydrolyse l'ATP et convertit cette énergie en déplacement relatif le long du microtubule orienté vers l’extrémité (-)
b. Dans le contexte des appendices subdistaux, où la dynéine est contrainte spatialement par la plateforme d’ancrage, ce mouvement ne sert pas au transport d’un cargo, mais se traduit par une force de traction centripète appliquée au microtubule ( cf. figure 1 ci-contre).
Cette traction plaque le microtubule contre l’appendice subdistal en augmentant la composante normale de la force entre le microtubule et la plateforme, ce qui stabilise le contact latéral avec le lattice proximal.
Le microtubule est ainsi maintenu dans une position proche du centrosome malgré les forces opposées du cytoplasme et du réseau d’actine qui exerce des forces tangentielles et centrifuges sur les microtubules, en particulier sur leurs portions distales, qui se transmettent mécaniquement jusqu’à leur extrémité (-).
c. En parallèle, la traction empêche la diffusion latérale et la rotation libre du microtubule en imposant une contrainte mécanique continue qui réduit ses degrés de liberté.
Le microtubule ne peut plus glisser ni pivoter autour de son axe, car tout déplacement latéral induit un désalignement immédiatement compensé par la force exercée par le moteur.
Remarque : l’activité motrice dirigée vers l’extrémité moins de la dynéine pourrait aussi :
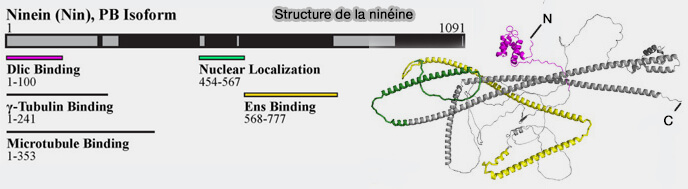
ancrer des complexes γ-tubuline en l’absence de dynéine/dynactine, potentiellement via une région de liaison composite des acides aminés 1-246 et 1179-1931 qui pourrait permettre l’ancrage direct des extrémités (-) des microtubules ( cf. figure 2),
déplacer les cylindres centriolaires le long des microtubules émanant de chaque centriole opposé, rapprochant ainsi les centrioles mère et fille ( cf. figure 3).
3. Ce mécanisme permet :
le maintien d’un réseau microtubulaire radial cohérent centré sur le centrosome,
l’absorption et la redistribution des contraintes mécaniques cytoplasmiques exercées sur les microtubules,
le positionnement stable du centrosome en interphase,
la préparation correcte de l’orientation du fuseau lors de l’entrée en mitose.
La perte de la dynéine, de Ndel1/Lis1 ou de la ninéine entraîne une désorganisation radiale du réseau, malgré une nucléation intacte.
MAP associées au centrosome
Les MAP (Microtubule-Associated Proteins) sont recrutées transitoirement autour du centrosome et remplissent plusieurs fonctions complémentaires :
stabiliser les microtubules en cours d’assemblage, en se liant aux segments naissants pour limiter leur dépolymérisation et assurer une croissance régulière, comme par exemple MAP7/ensconsin.
renforcer les asters mitotiques, principalement assurés par les MAP organisatrices, en consolidant les interactions latérales entre microtubules, ce qui accroît la rigidité et la cohésion du réseau astral, tel que NuMA, qui organise les microtubules astraux autour du pôle mitotique,
moduler la dynamique microtubulaire en permettant des ajustements rapides du réseau en réponse aux forces exercées lors de la formation du fuseau, comme CLASP1, qui stabilise les extrémités et prévient la catastrophe.






 MAP associées au centrosome).
MAP associées au centrosome).