La majorité des +TIPs ne reconnaissent pas directement l’extrémité (+) des microtubules, mais sont recrutées via les protéines EB par des mécanismes moléculaires spécialisés.
La plupart des +TIPs ne reconnaissent pas directement la géométrie de l’extrémité (+) des microtubules, mais sont recrutées secondairement via les protéines EB, qui jouent le rôle de plateforme centrale d’assemblage.
Ce recrutement repose principalement sur deux types d’interactions moléculaires, impliquant soit des motifs SxIP, soit des domaines CAP-Gly, permettant une association hiérarchisée et dynamique des +TIPs à EB1/EB3.
Cette organisation EB-dépendante génère une plateforme directionnelle transitoire, capable d’orienter la croissance des microtubules, de coordonner leur capture corticale et de moduler localement la stabilité de la coiffe GTP.
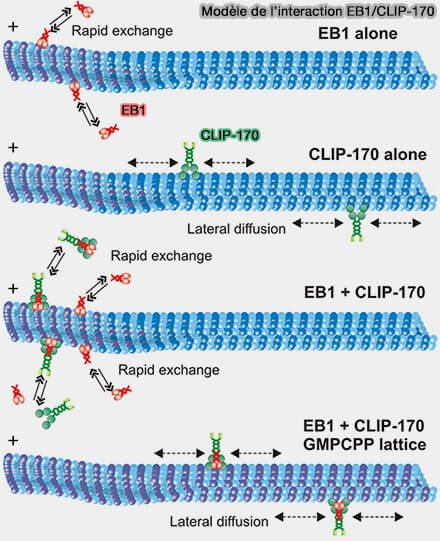
EB1 et CLIP-170
(Figure : vetopsy.fr d'après Chen et coll)
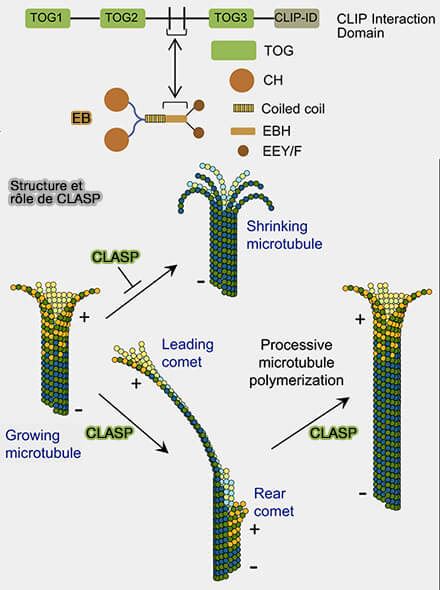
1. CLIP-170 est recruté aux extrémités (+) et contient :
deux domaines CAP-Gly N-terminaux, notés A et B,, flanqués par des régions riches en sérine, et plusieurs motifs SXIP et SXIP-like responsables de la reconnaissance des motifs EEY portés par EB1 et certaines tubulines modifiées,
une région coiled-coil centrale impliquée dans la dimérisation de la protéine ;
deux motifs zinc knuckles (CCHC) situés dans la région C-terminale, coordonnant des ions zinc et contribuant à la stabilisation structurale de cette portion de la protéine, impliqués :
dans la régulation de son interaction intramoléculaire, limitant l’accès du domaine CAP-Gly aux motifs EEY des +TIPs et des tubulines modifiées, ce qui module finement la capacité de capture des extrémités (+) et l’association dynamique avec les partenaires tels qu’EB1
dans les interactions avec des effecteurs tels que p150Glued et LIS1, renforçant le couplage entre les extrémités (+) des microtubules et les complexes moteurs ou régulateurs.
Interaction EB1/CLIP-70
(Figure : vetopsy.fr d'après Dixit et coll)un motif C-terminal de type EEY/ETF, capable d’interagir avec les domaines CAP-Gly dans un mode similaire aux motifs EEY portés par EB1, participant à l’organisation multivalente et régulée du complexe CLIP-170-EB1.
2. L’interaction avec EB1 positionne CLIP-170 à la frontière entre le lattice GTP-like du microtubule en croissance et la zone distale de l’extrémité (+)
Cette organisation permet à CLIP-170 de relier la dynamique de l’extrémité (+) aux structures corticales et aux complexes moteurs, assurant à la fois :
de participer à l’ancrage cortical transitoire lors des phases de ralentissement ou de reconnaissance spatiale,
Elles sont traitées ici en tant que +TIPs car leur activité est spatialement ciblée par un recrutement EB-dépendant au voisinage des extrémités (+) en croissance, ce qui en fait un exemple central pour comprendre comment la plateforme EB positionne des fonctions stabilisatrices locales sans intervenir directement dans l’élongation.
Remarque : les CLASP n'ont rien à voir avec les protéines CLASP (CLathrin Associated Sorting Proteins ou protéines de tri associées à la clathrine) aui désignent les protéines qui lient le cargo dans le puits recouvert de clathrine (CCP)
1. CLASP1/2 agissent en détectant les distorsions locales du lattice et les géométries défavorables à la croissance, correspondant à des régions où les protofilaments ne sont plus parfaitement alignés ni latéralement appariés.
Structure et rôle de CLASP
(Figure : vetopsy.fr d'après Aher et coll)
a. Moléculairement, ces distorsions se manifestent par :
une courbure modérée des protofilaments,
une perte partielle des contacts latéraux entre protofilaments adjacents,
une transition locale du lattice GTP-like vers une géométrie moins stable.
b. Les domaines TOG (Tumor Overexpressed Gene) de CLASP présentent une affinité préférentielle pour ces états intermédiaires de la tubuline polymérisée, qu’ils reconnaissent par leur géométrie plutôt que par une séquence ou un état nucléotidique strict (CLASP Suppresses Microtubule Catastrophes through a Single TOG Domain 2018).
2. Cette reconnaissance permet à CLASP de se concentrer précisément sur les zones susceptibles d’entrer en catastrophe :
empêchant la propagation des protofilaments ouverts ou désynchronisés,
Remarque : CLASP exerce ainsi une activité stabilisatrice du microtubule, ce qui justifie sa présence dans les tableaux des MAP stabilisatrices, bien que son ancrage principal soit traité dans le cadre des +TIPs.
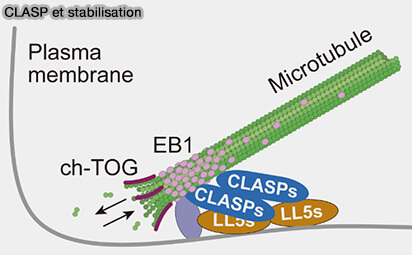
3. Au cortex cellulaire, CLASP1/2 s’associent au lattice proximal des microtubules en croissance et sont recrutées sur des plateformes membranaires telles que LL5α/β, ce qui relocalise l’activité de stabilisation décrite ci-dessus dans un contexte cortical spatialement contraint.
Cette relocalisation repose sur son domaine CLIC (CLASP Interaction with the Cortex), qui assure le ciblage cortical de CLASP en médiant ses interactions avec des plateformes membranaires riches en phosphoinositides.
I
CLASP et stabilisation
(Figure : vetopsy.fr d'après Nakamura et coll)
Remarque : LL5α/β sont des adaptateurs corticaux associés aux phosphoinositides membranaires, qui servent de plateformes d’ancrage pour les +TIPs et contribuent à la capture et à l’organisation des microtubules à l’interface cortex-membrane.
Couplage CLIP-170/CLASP
Le module CLIP-170/CLASP assure ainsi une continuité fonctionnelle entre :
la capture et l’ancrage des extrémités (+) par CLIP-170,
la stabilisation dynamique locale et la prévention des catastrophes par CLASP.
Ce couplage est particulièrement important au cortex cellulaire et dans le fuseau mitotique, où la robustesse des microtubules repose davantage sur le contrôle fin des transitions croissance/pause/dépolymérisation que sur la seule vitesse d’élongation.
APC et spectraplakines (ACF7/MACF1)
APC (Adenomatous Polyposis Coli) et les spectraplakines, telles qu’ACF7/MACF1, sont des +TIPs spécialisés dans le guidage directionnel et l’ancrage cortical des microtubules, en assurant le couplage fonctionnel entre microtubules, cytosquelette d’actine et structures d’adhérence.
1. APC agit comme un facteur de guidage des extrémités (+) en croissance.
a. APC est recrutée aux extrémités (+) des microtubules via des motifs SxIP reconnus par le domaine EBH des protéines EB.
Ce recrutement permet à APC d’orienter préférentiellement la croissance des microtubules vers des régions corticales spécifiques, notamment les sites d’adhérence et les domaines de polarité cellulaire.
b. APC ne stabilise pas directement la dynamique microtubulaire, mais impose une directionnalité spatiale à la croissance en reliant la plateforme EB à des signaux corticaux et actiniques.
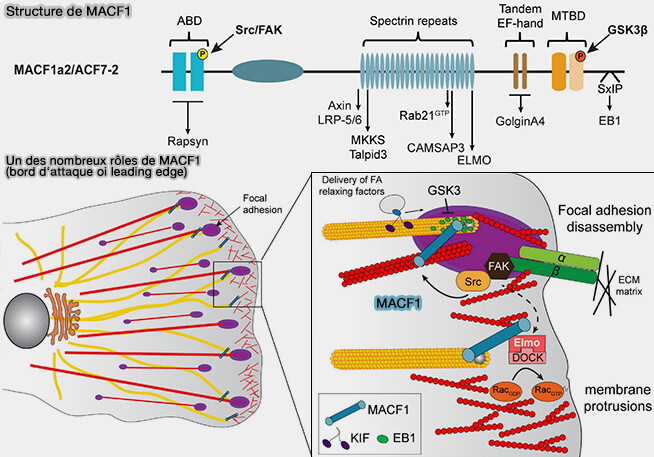
2. Les spectraplakines, i.e. ACF7/MACF1 (Actin Crosslinking Factor 7/Microtubule Actin Crosslinking Factor 1) assurent l’ancrage mécanique cortex-microtubules, mais ce rôle est principalement mécanique et organisationnel, distinct de la régulation fine de la dynamique exercée par CLASP ou XMAP215/ch-TOG (A Multifaceted Giant Protein Microtubule-Actin Cross-Linking Factor 1 2025).
ACF7/MACF1 sont de grandes protéines de pont, MACF1/ACF7 appartient à la famille des plakines, capables de lier simultanément les microtubules, les filaments d’actine et des complexes d’adhérence.
Elles stabilisent durablement les contacts entre microtubules corticaux et cortex d’actine, augmentant la persistance des microtubules orientés et limitant leur réorientation aléatoire.
L’interaction EB-dépendante d’APC initie un guidage directionnel des microtubules, tandis que les spectraplakines convertissent ce guidage en ancrage stable et persistant au cortex.
Rôle de MACF1 au bord d‘attaque (leading edge)
(Figure : vetopsy.fr d'après Cusseaddu et coll)
Certaines +TIPs influencent directement la cinétique d’élongation en stabilisant les protofilaments naissants ou en modifiant la géométrie de l’extrémité.
XMAP215/ch-TOG
1. XMAP215/ch-TOG agit comme un polymérase de la tubuline à l’extrémité (+).
Son nom dérive respectivement de Xenopus microtubule-associated protein of 215 kDa et de colonic and hepatic tumor overexpressed gene.
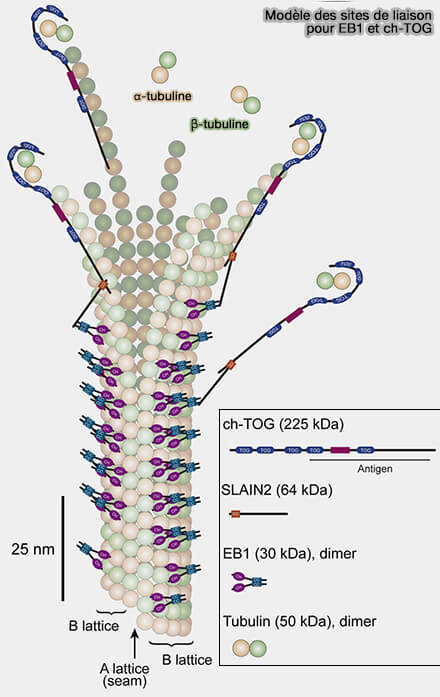
Modèle des sites de liaison pour EB1 et ch-TOG
(Figure : vetopsy.fr d'après Nakamura et coll)
b. Ce mécanisme permet :
l’alignement correct des dimères avec les protofilaments existants,
la réduction de la barrière énergétique d’ajout d’un nouveau dimère,
la synchronisation de la croissance entre protofilaments voisins.
Par ce mécanisme, XMAP215/ch-TOG allonge activement les protofilaments, ce qui stabilise indirectement la coiffe GTP en limitant l’exposition de protofilaments courts ou incomplets, principaux déclencheurs de catastrophe.
c. Il exerce cette fonction en :
stabilisant les embryons de protofilaments à l’extrémité (+),
augmentant fortement la vitesse d’incorporation de la tubuline-GTP,
favorisant l’allongement processif des protofilaments, même dans des conditions limitantes en tubuline libre.
2. XMAP215/ch-TOG coopère étroitement avec EB1, +TIP emblématique de la coiffe GTP.
EB1 reconnaît la conformation GTP-like de la tubuline et stabilise la géométrie de l’extrémité (+).
SLAIN2 par ses motifs SxIP-like (et régions riches en sérines) est recruté sur le lattice proximal par interaction avec le domaine EBH (EB homology domain) de EB1 où il agit comme adaptateur moléculaire.
SLAIN2 concourt, via une région centrale de SLAIN2 (non SxIP), , au recrutement de ch-TOG. et établit un lien direct entre EB1 et XMAP215/ch-TOG, augmentant la concentration locale de la polymérase.
une réduction de la probabilité de catastrophe pendant les phases d’élongation active.
b. Ce couplage est particulièrement critique dans les neurones (croissance axonale, plasticité) et dans le fuseau mitotique, où une croissance soutenue des microtubules est requise.
3. En outre, XMAP215/ch-TOG forme un complexe fonctionnel avec TACC3 après la phosphorylation mitotique de cette dernière par Aurora A, ce qui permet de cibler et de renforcer l’activité polymérase de XMAP215/ch-TOG sur les microtubules du fuseau.
1. NAV1 est recrutée aux extrémités (+) en croissance via la plateforme EB-dépendante, ce qui lui permet de s’accumuler dans les régions de croissance neuritique active.
a. Elle contribue à la progression des neurites précoces en couplant la dynamique microtubulaire à des signaux corticaux locaux, favorisant une croissance orientée plutôt qu’aléatoire.
Modèle du rôle de NAV1 dans la dynamique des cônes de coissance
(Figure : vetopsy.fr d'après Sánchez-Huertas et coll)
b. NAV1 agit comme intégrateur fonctionnel qui permet une coordination fine entre extension microtubulaire et remodelage cortical par :
la dynamique des microtubules,
les forces de traction dépendantes de l’actomyosine.
2. Ainsi, NAV1 ne modifie pas directement la cinétique d’élongation, mais impose un biais directionnel stable, essentiel à l’organisation spatiale des neurites en formation.
+TIPs spécialisés
Certaines +TIPs assurent des fonctions ciblées, associées à des structures cellulaires spécifiques ou à des contextes particuliers du cycle cellulaire et de la différenciation.
1. Les kinésines-8 (KIF18A, KIF18B) sont des moteurs associés aux extrémités (+) des microtubules, spécialisés dans la régulation de leur longueur et de leur dynamique, en particulier dans le contexte mitotique.
Elles ne participent pas à la nucléation des microtubules, mais modulent leur extension et leur persistance en fonction de leur localisation et des contraintes mécaniques du fuseau mitotique.
a. KIF18A est principalement exploitée au cours de la mitose pour le contrôle fin des microtubules kinétochoriens.
Elle s’accumule préférentiellement à proximité de la plaque équatoriale et limite les fluctuations excessives des extrémités (+) des microtubules kinétochoriens.
Son activité est étroitement coordonnée par des MAP périchromosomiques, notamment HURP, avec lesquelles elle ajuste la dynamique microtubulaire afin de stabiliser l’alignement métaphasique des chromosomes ( HURP et kif 18A).
Associée aux extrémités (+) de manière EB-dépendante, elle régule la longueur des microtubules périphériques du fuseau et contribue à son positionnement et à son centrage dans la cellule.
KIF18B participe ainsi à l’ajustement global de la géométrie du fuseau mitotique, sans intervenir directement dans l’alignement chromosomique.
2. SLAIN1/2, principalement impliquées dans le développement neuronal, agissent comme adaptateurs EB-dépendants favorisant une croissance microtubulaire soutenue.
Elles servent de relais entre EB1 et XMAP215 pour soutenir une croissance axonale soutenue ( interactions)
en participant au positionnement nucléaire et en organisant les forces intracellulaires, notamment dans les cellules migrantes et les neurones en développement.





 I
I


 HURP et kif 18A).
HURP et kif 18A).