


Noyau
Pores nucléaires : vue d'ensemble et architecture
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

Les pores nucléaires sont des complexes macromoléculaires insérés dans l’enveloppe nucléaire, organisés en une charpente protéique annulaire ancrée dans la double membrane, qui assurent les échanges sélectifs entre noyau et cytoplasme.
Le complexe du pore nucléaire (NPC) est une structure multiprotéique insérée dans l’enveloppe nucléaire et assurant les échanges sélectifs entre noyau et cytoplasme ( transport nucléocytoplasmique).
transport nucléocytoplasmique).
- Il traverse la double membrane, externe et interne, au niveau de leur zone de fusion et forme un canal central entouré d’une charpente annulaire organisée selon une symétrie octogonale.
- La membrane nucléaire externe, dans laquelle s’insère le pore, demeure en continuité topologique avec le réticulum endoplasmique (RE), assurant l’intégration du complexe du pore nucléaire au système endomembranaire global de la cellule.
Au-delà de sa fonction de transport, le pore nucléaire constitue un élément structural majeur de l’enveloppe nucléaire, participant à son organisation mécanique et à son intégration dans l’architecture globale du noyau.
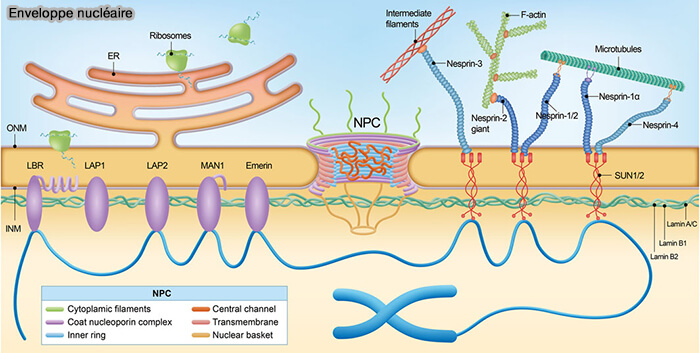
(Figure : vetopsy.fr d'après Wu et coll)
Organisation architecturale du complexe du pore nucléaire (NPC)
Le complexe du pore nucléaire (NPC) forme ainsi un cylindre protéique inséré dans la double membrane nucléaire, dont la hauteur est d’environ 60 à 80 nm et le diamètre externe d’environ 100 à 120 nm chez les mammifères.
Une cellule somatique de vertébré contient en moyenne 2 000 à 3 000 pores nucléaires, ce nombre variant selon le type cellulaire, la taille nucléaire et l’intensité des échanges nucléocytoplasmiques, soulignant l’importance quantitative et fonctionnelle de ces complexes.
1. Le NPC est un assemblage macromoléculaire d’environ 100-120 MDa chez les vertébrés, constitué d’environ 30 types de nucléoporines (Nups) organisées en multiples copies qui présente une symétrie radiale octogonale autour d’un axe central perpendiculaire au plan de l’enveloppe nucléaire.
- La symétrie octogonale ne correspond pas à huit couches empilées verticalement, mais à huit unités répétitives disposées radialement autour du canal central, visibles en vue de dessus.
- En coupe longitudinale, ces unités fusionnent visuellement pour former des anneaux cylindriques continus.
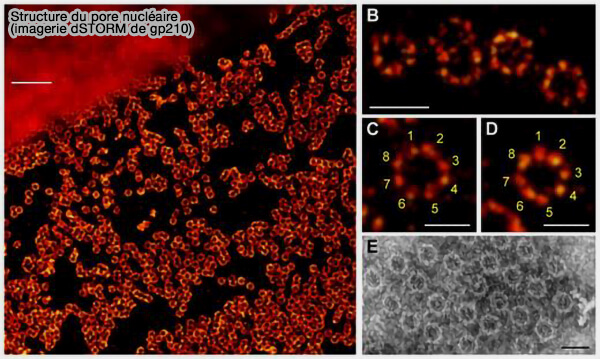
(Figure : vetopsy.fr d'après Yang et coll)
2. La symétrie octogonale qui en résulte se manifeste par un réseau de sous-complexes structuraux formant (Super-resolution imaging visualizes the eightfold symmetry of gp210 proteins around the nuclear pore complex and resolves the central channel with nanometer resolution 2012) :
- l’anneau interne, qui constitue le cœur structural du complexe et stabilise la courbure membranaire au niveau de la zone de fusion,
- l’anneau cytoplasmique,
- l’anneau nucléaire.
3. Un canal central traverse la zone de fusion des membranes interne et externe et constitue la voie de transport nucléocytoplasmique.
Le diamètre fonctionnel du canal central est d’environ 40 à 50 nm et sa dimension axiale varie généralement entre 60 et 80 nm, ces valeurs pouvant fluctuer selon l’état conformationnel et la méthode d’analyse structurale.
Charpente protéique centrale du pore (scaffold)
Anneau interne
1. L’anneau interne constitue le noyau architectural du pore nucléaire et occupe la position médiane du complexe, exactement à l’endroit où les membranes nucléaires externe et interne fusionnent.
- C’est lui qui stabilise la forte courbure membranaire imposée par la jonction des deux membranes.
- Il correspond à la masse protéique centrale située entre l’anneau cytoplasmique (au-dessus) et l’anneau nucléaire (en dessous).
1. Il forme un cylindre protéique épais qui entoure directement le canal central et en délimite le diamètre fonctionnel.
- Il sert de plateforme d’ancrage aux nucléoporines riches en répétitions FG, dont les domaines intrinsèquement désordonnés s’étendent vers l’intérieur du canal pour former la barrière sélective.
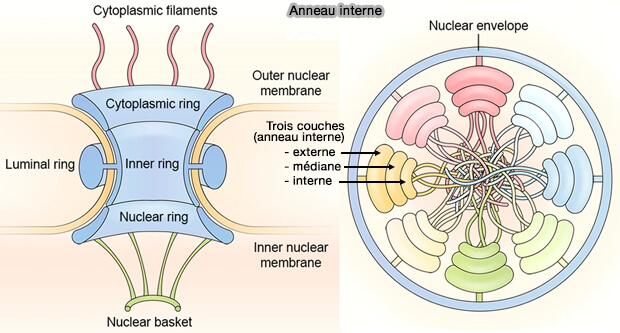
(Figure : vetopsy.fr d'après Yang et coll)
2. Les sous-complexes répétés qui composent l’anneau interne sont disposés selon les huit rayons de la symétrie octogonale.
a. Dans certaines reconstructions cryo-EM récentes, l’anneau interne apparaît organisé en trois couches concentriques radiales appartenant à une même unité annulaire (Nuclear transport proteins: structure, function and disease relevance 2023)
- La couche la plus interne, située au contact du canal central, correspond aux nucléoporines qui ancrent les domaines FG et participent à la constitution de la barrière sélective.
- La couche médiane, qui constitue le noyau structural de l’anneau interne, est formée de nucléoporines structurales allongées, complexe en Y, qui assurent la rigidité mécanique du cylindre central.
- La couche externe est positionnée au voisinage immédiat de la membrane incurvée et établit des interactions avec des nucléoporines transmembranaires et assure la continuité entre l’anneau interne et l’enveloppe nucléaire.
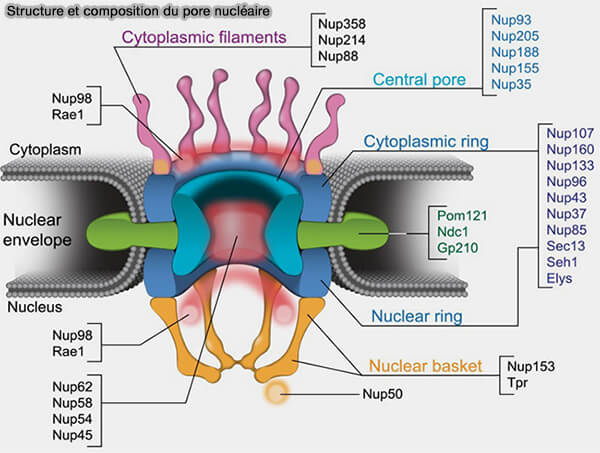
(Figure : vetopsy.fr d'après Ibarra et Hetzer)
b. En outre, l’anneau interne des vertébrés présente une organisation stratifiée le long de l’axe nucléocytoplasmique en deux sous-couches superposées, correspondant aux composantes cytoplasmique et nucléaire de l'anneau interne.
Cette organisation axiale explique la présence de deux cylindres centraux distincts dans les représentations en coupe longitudinale, sur la figure eux-mêmes divisés en deux.
3. Chaque module radial ne constitue pas un bloc isolé, mais un sous-complexe possédant des interfaces latérales spécifiques permettant son association avec les modules adjacents.
a. Les huit unités répétitives s’imbriquent ainsi par des interactions protéine–protéine complémentaires pour former un anneau fermé continu, sans discontinuité structurale ( complexes en Y).
b. L’assemblage circulaire résulte d’un réseau polygonal dans lequel les domaines allongés des sous-complexes s’interconnectent latéralement, tandis que la courbure imposée par la fusion membranaire favorise leur disposition en symétrie radiale.
Cette organisation en treillis annulaire confère au complexe une rigidité mécanique élevée tout en conservant une architecture modulaire compatible avec une biogenèse progressive.
Anneaux externes
Ces deux anneaux constituent les couches périphériques du complexe du pore nucléaire et encadrent axialement l’anneau interne.
Ils assurent la continuité architecturale entre la charpente centrale et les extensions périphériques dynamiques, tout en participant à la stabilisation mécanique globale du complexe (The cellular environment shapes the nuclear pore complex architecture 2021).
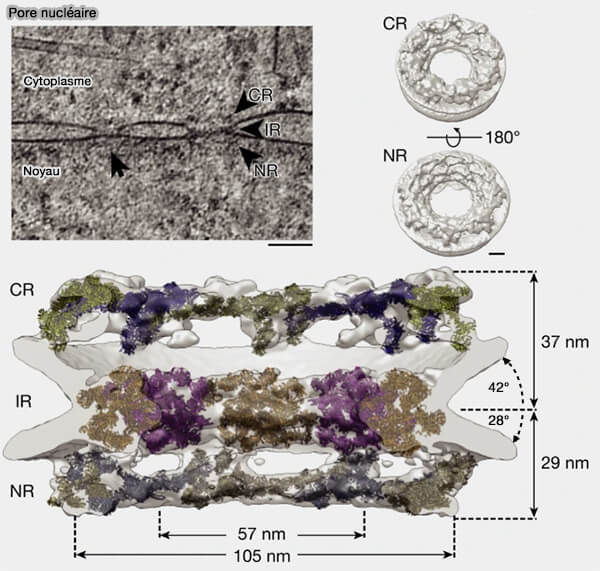
(Figure : vetopsy.fr d'après Schuller et coll)
Anneau cytoplasmique
1. L’anneau cytoplasmique est situé du côté cytosolique du pore nucléaire et correspond, en coupe longitudinale, à la couche supérieure du complexe.
- Il est disposé immédiatement au-dessus de l’anneau interne et contribue à l’élargissement périphérique du cylindre structural.
- Sur le plan structural, l’anneau cytoplasmique conserve la symétrie octogonale du complexe et s’organise en unités répétitives alignées avec celles de l’anneau interne, assurant ainsi la continuité radiale du pore.
2. L’anneau cytoplasmique :
- sert de point d’ancrage aux filaments cytoplasmiques,
- constitue la face d’entrée et de sortie du canal central et participe aux premières interactions des complexes de transport (transport nucléocytoplasmique).
Anneau nucléaire
L’anneau nucléaire est situé du côté nucléoplasmique du complexe du pore nucléaire et correspond, en coupe longitudinale, à la couche inférieure du cylindre structural.
- Il est disposé immédiatement sous l’anneau interne et constitue la principale interface entre le canal central et le compartiment nucléaire.
- Sur le plan structural, l’anneau nucléaire conserve la symétrie octogonale du complexe et s’organise en unités répétitives alignées avec celles de l’anneau interne et de l’anneau cytoplasmique, assurant la continuité géométrique du pore.
1. L’anneau nucléaire constitue une interface structurale bidirectionnelle entre le canal central et le nucléoplasme.
- Il sert de point d’ancrage au panier nucléaire, extension périphérique projetée dans le nucléoplasme.
- Il établit des interactions transitoires avec les complexes d’import et d’export (transport nucléocytoplasmique).
2. L’anneau nucléaire est positionné au contact de la chromatine périphérique.
a. Certaines régions génomiques, notamment des domaines transcriptionnellement actifs, peuvent s’associer de manière préférentielle à des pores nucléaires.
Des gènes fortement exprimés peuvent être recrutés à proximité du pore, ce qui favorise un couplage spatial entre transcription nucléaire et export des ARN nouvellement synthétisés.
b. Cette proximité contribue :
- à l’organisation locale de la chromatine, en participant à la structuration de microdomaines nucléaires où se concentrent certaines activités transcriptionnelles,
- à la coordination entre transcription et export des ARN, en réduisant la distance physique entre le site de synthèse de l’ARN messager et le canal d’export,
- au contrôle qualité des ARN messagers avant leur libération dans le cytoplasme, certaines étapes de maturation et de surveillance étant associées à la face nucléaire du pore avant la libération définitive des transcrits dans le cytoplasme.
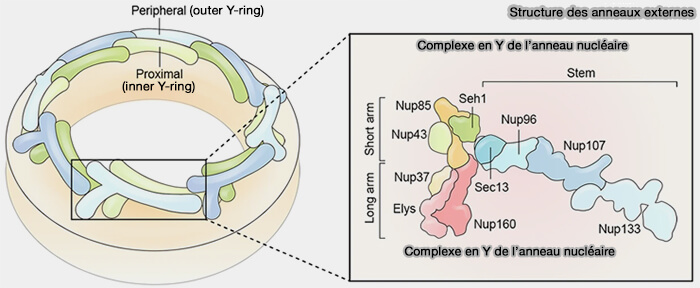
(Figure : vetopsy.fr d'après Yang et coll)
Ossature des anneaux externes
1. Le complexe en Y constitue le module structural majeur des anneaux cytoplasmique et nucléaire du complexe du pore nucléaire.
- Huit unités répétitives s’assemblent radialement pour former un anneau continu participant à la symétrie octogonale du pore.
2. Il forme l’ossature des anneaux externes du pore nucléaire et encadre le canal central sans participer directement à la barrière sélective.
Insertion dans la membrane
1. Les nucléoporines transmembranaires sont insérées dans la zone de fusion entre la membrane nucléaire interne et la membrane nucléaire externe.
- Contrairement aux complexes structuraux qui constituent les anneaux du pore, les nucléoporines transmembranaires ne forment pas une structure annulaire autonome.
- Elles assurent le couplage mécanique entre la bicouche lipidique et la charpente protéique du complexe.
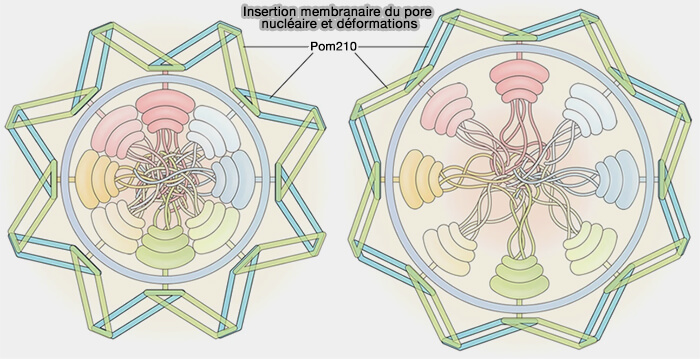
(Figure : vetopsy.fr d'après Yang et coll)
2. Dans l’espace périnucléaire, au niveau de la zone de fusion, certaines études structurales récentes décrivent la présence d’un luminal ring, correspondant à un épaississement annulaire associé aux nucléoporines transmembranaires (Nuclear transport proteins: structure, function and disease relevance 2023)
Cette structure périnucléaire entoure l’orifice du pore du côté luminal et contribue à la stabilisation de l’interface membrane–pore sans constituer un anneau protéique interne du NPC.
3. En reliant les anneaux externes et la périphérie de l’anneau interne à la membrane nucléaire, les nucléoporines transmembranaires maintiennent la cohésion mécanique du complexe et stabilisent la courbure membranaire imposée par l’insertion du pore.
Suite de la structure et organisation fonctionnelle ey dynamique
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauEnveloppe nucléairePores nucléairesLaminaCorps nucléairesTransport nucléocytoplasmiqueCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteFilaments d'actineFilaments intermédiairesMicrotubulesReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation