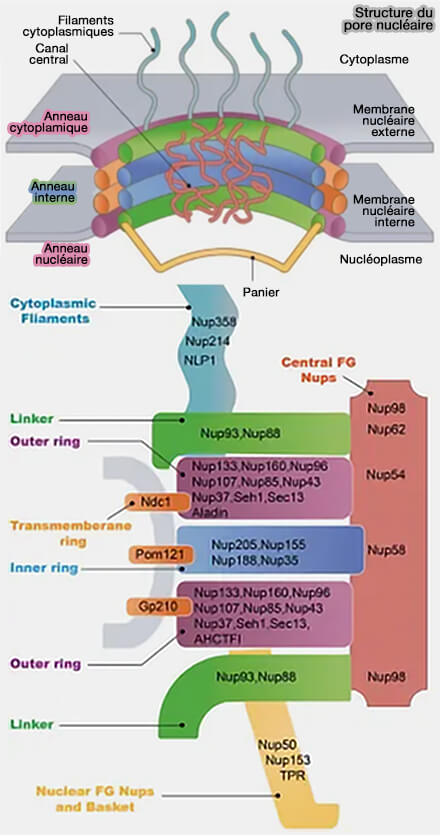
Les nucléoporines sont les sous-unités protéiques du complexe du pore nucléaire, et les nucléoporines annulaires en organisent les anneaux structuraux assurant l’architecture du canal de transport.
Chez les vertébrés, environ trente types distincts de nucléoporines sont identifiés.
Assemblées en multiples copies, elles forment un complexe macromoléculaire d’environ 100–120 MDa inséré dans l’enveloppe nucléaire.
2. Les nucléoporines ne sont pas réparties de manière uniforme, mais organisées en sous-complexes structuraux hiérarchisés correspondant aux différents modules du pore :
3. Cette organisation modulaire reflète une spécialisation structurale des nucléoporines :
certaines assurent la rigidité mécanique du pore.
d’autres confèrent au canal sa propriété sélective,
d’autres encore participent à l’intégration membranaire et nucléaire.
Nucléoporines annulaires
Les nucléoporines structurales annulaires constituent la charpente rigide du complexe du pore nucléaire, assurant la stabilité mécanique du canal de transport en participant à la formation de :
Ces nucléoporines assurent l’assemblage radial et la stabilité à long terme de la charpente du pore.
Nucléoporines des anneaux externes : complexe en Y
Le complexe en Y, appelé Nup107-160,constitue le module central de la charpente externe du complexe du pore nucléaire des anneaux cytoplasmique et nucléaire( charpente du pore).
Structure et nucléoporines du pore nucléaire
(Figure : vetopsy.fr d'après Wu et coll)
Il forme l’ossature principale des anneaux externes et encadre le canal central sans participer directement à la barrière FG.
Il sert également de plateforme structurale pour l’ancrage d’autres nucléoporines impliquées dans l’architecture périphérique du NPC.
Remarque : Il est souvent appelé Nup107-160, du fait de deux de ces nucléoporines principales, car il regroupe notamment, chez les vertébrés, Nup107, Nup160, Nup133, Nup96, Nup85, Nup43, Sec13 et Seh1
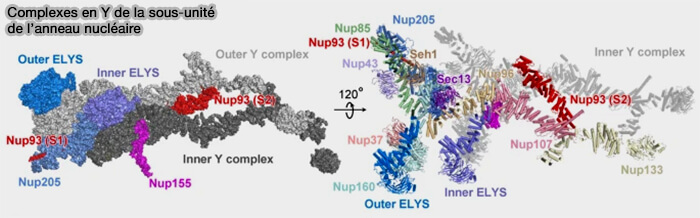
1. Le complexe adopte une architecture tridimensionnelle en forme de Y, composée d’un tronc central et de deux bras divergents.
a. Huit copies s’assemblent radialement pour former un anneau continu.
b. Chez les vertébrés, chaque anneau externe comprend deux couches coaxiales de complexes en Y, dites périphérique (outer Y-ring) et proximale (inner Y-ring), organisées axialement au sein des anneaux cytoplasmique et nucléaire.
Nup93 et Nup205, constituants de l’anneau interne, assurent le pontage axial entre ces deux couches de complexes en Y sans en constituer des éléments répétitifs.
2. L’association de modules α-solénoïdes allongés et de plateformes β-propeller compactes permet l’assemblage d’un réseau polygonal stable tout en conservant une organisation modulaire.
a. Le tronc central du complexe est formé principalement par Nup107 et Nup96, assurant la continuité structurale entre les deux bras et la cohésion du module.
b. Les bras allongés du complexe confèrent aux nucléoporines une grande surface d’interaction latérale et une certaine flexibilité modulaire.
Nup160 constitue l’élément majeur d’un des bras longs du complexe, formant une portion allongée participant à l’extension périphérique du module.
Nup85, associé à Seh1, participe à la formation de l’autre bras divergent du Y, contribuant à l’interface avec les complexes voisins dans l’anneau externe.
Complexe en Y de la sous-unité de l'anneau nucléaire
(Figure : vetopsy.fr d'après Huang et coll)
4. Les domaines β-propeller sont formés par l'arrangement de quatre à huit feuillets β (β-sheet) en forme de " lame " autour d'un axe central selon une configuration toroïdale, fréquemment utilisés dans les plateformes d’interaction protéine–protéine (solénoïdes circulaires).
Ces domaines sont retrouvés dans :
Nup133, dont le domaine N-terminal adopte une architecture β-propeller positionnée à l’extrémité d’un bras du complexe, intervenant comme plateforme d’interaction protéin--protéine et de point de connexion avec les modules α-solénoïdes adjacents,
Sec13 et Seh1, sous-unités adaptatrices compactes intégrées à la charpente externe, participant à la stabilisation latérale du réseau polygonal formé par les complexes en Y,
Nup43, contribuant aux interfaces d’assemblage et à la cohésion structurale du module externe.
5. On trouve aussi deux nucléoporines associées au complexe en Y, mais qui n'en constituent pas un élément répétitif.
a. ALADIN (AAAS), dans l'anneau cytoplasmique, contribue à la stabilisation périphérique du pore et participe aux interfaces avec les filaments cytoplasmiques et certaines nucléoporines FG du côté cytosolique.
Elle s'associe avec le bras long du complexe en Y.
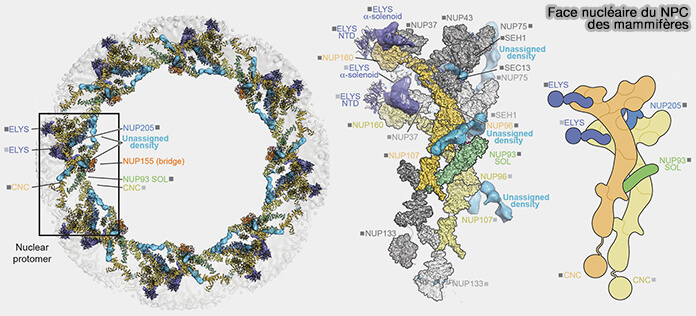
Dans la figure ci-dessus, les mentions " outer ELYS " et " inner ELYS " ne correspondent pas à deux formes distinctes d’ELYS, mais à des positions axiales différentes d’une même molécule associée aux complexes en Y de la couche périphérique (outer Y-ring) de l’anneau nucléaire, dont la densité peut apparaître projetée vers la couche proximale (inner Y-ring).
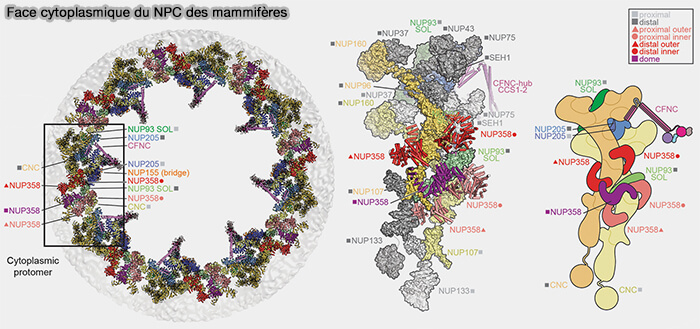
Face cytoplasmique du pore
(Figure : vetopsy.fr d'après Bley et coll)
Nup93 agit comme élément organisateur central du module interne et participe aux interfaces de liaison avec les complexes en Y des anneaux externes via ses interactions avec Nup205 et Nup188.
Nup205 et Nup188, grandes nucléoporines à repliement α-solénoïde, forment des segments allongés du cylindre central dont certaines surfaces latérales établissent des interfaces de contact avec les couches périphérique et proximale de complexes en Y des anneaux externes.
Nup155, établit la connexion entre l’anneau interne et les complexes en Y des anneaux externes en assurant l’ancrage de ces derniers au scaffold interne, contribuant ainsi à la continuité structurale entre les modules interne et externe du NPC.
Remarque : contrairement aux autres nucléoporines du module interne, Nup35 ne constitue pas un élément transmembranaire du NPC.
Dépourvue de domaine transmembranaire, elle agit comme une nucléoporine périphérique associée à la membrane nucléaire interne grâce à des hélices amphipathiques capables de s’y ancrer de manière transitoire.
Par ses interactions avec Nup93 et certaines nucléoporines transmembranaires telles que Ndc1, Nup35 assure un couplage mécanique adaptable entre la charpente interne du pore et la membrane nucléaire, ses régions flexibles facilitant l’ajustement local à la courbure membranaire au niveau du pore.
Face nucléaire du pore
(Figure : vetopsy.fr d'après Bley et coll)
2. Sur le plan structural, le module interne est dominé par des architectures de type α-solénoïde allongées organisées en répétitions radiales, permettant :
la stabilisation du diamètre du canal central,
le maintien de la symétrie radiale,
la transmission des contraintes mécaniques entre membrane et charpente du pore.
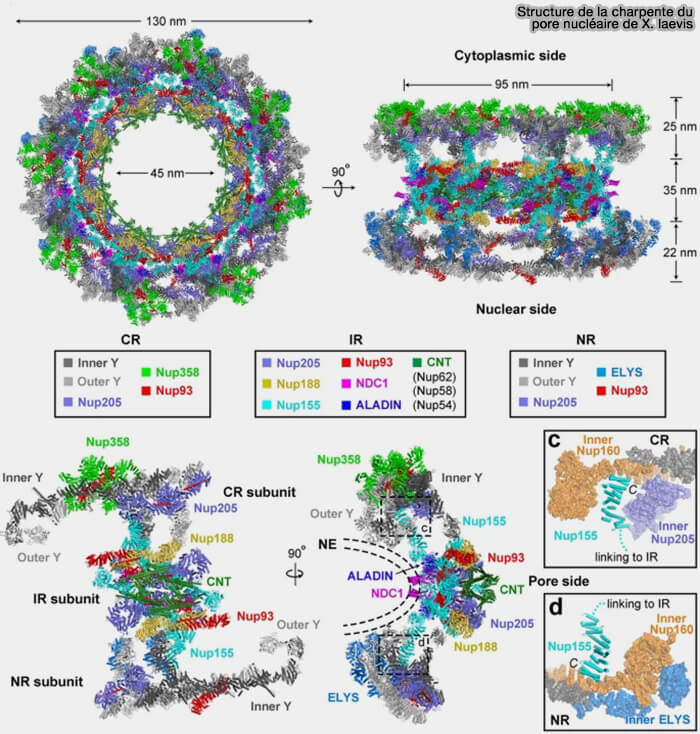
Structure de la charpente du pore nucléaire de X. laevis
(Figure : vetopsy.fr d'après Huang et coll)




 charpente du pore).
charpente du pore).