Le transport nucléocytoplasmique assure les échanges directionnels de macromolécules entre noyau et cytoplasme via le complexe du pore nucléaire, sous le contrôle du gradient de Ran.
Principes généraux du transport à travers le NPC
La compartimentation nucléaire impose un système d’échanges hautement contrôlé permettant de coordonner transcription, réplication et signalisation cellulaire (Structure and gating of the nuclear pore complex 2015).
1. La diffusion passive permet le passage bidirectionnel de petites molécules et de protéines de faible masse, généralement ≤ 20–30 kDa, seuil dépendant du rayon hydrodynamique et de la conformation, selon leur gradient de concentration.
Ce mécanisme est non saturable, indépendant du système Ran et ne requiert aucun signal de ciblage spécifique.
Par exemple, des ions (Ca++, K+), des nucléotides (ATP, GTP) ou de petites protéines cytosoliques comme l’ubiquitine (~8,5 kDa) peuvent diffuser librement à travers le pore nucléaire sans intervention de récepteurs de transport.
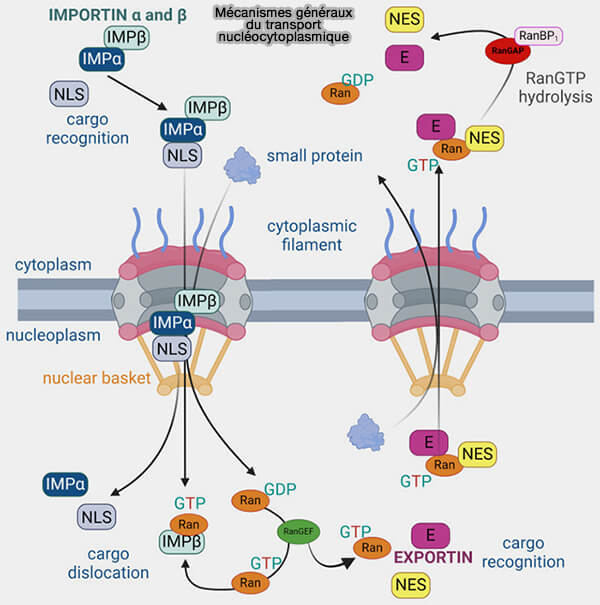
Mécanismes généraux du transport nucléocytoplasmique
(Figure : vetopsy.fr d'après Zaitsava et coll)
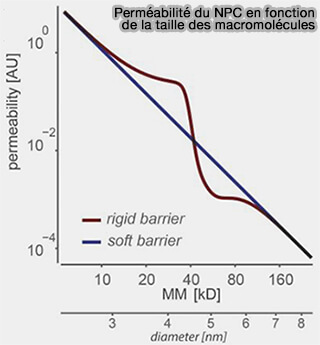
2. Contrairement au modèle historique de barrière rigide postulant un seuil de diffusion passif strict (~40–60 kDa), le complexe du pore nucléaire fonctionne comme une barrière souple (soft barrier) dans laquelle la perméabilité décroît progressivement avec l’augmentation du rayon hydrodynamique des macromolécules, dépendant notamment de leur compacité tridimensionnelle (Simple rules for passive diffusion through the nuclear pore complex 2016).
a. Une protéine globulaire compacte proche de 40 kDa peut encore diffuser lentement à travers le pore nucléaire, sans pour autant assurer un transport physiologiquement efficace.
Perméabilité du pore
(Figure : vetopsy.fr d'après Timney et coll)
Un complexe allongé ou multimérique de masse comparable, par exemple un dimère de facteur de transcription tel que NF-κB (p50/p65), possède un rayon hydrodynamique plus important, ce qui impose un transport médié par récepteurs.
b. Au-delà de ~50-60 kDa, la diffusion passive devient physiologiquement négligeable, comme pour p53 (~53 kDa), les polymérases nucléaires, les histones lorsqu’elles sont intégrées en complexes, ou encore les sous-unités ribosomiques.
3. Le mécanisme actif repose sur des déterminants moléculaires spécifiques et sur une organisation séquentielle du cycle de transport.
a. Le transport actif est un processus sélectif, saturable et vectoriel reposant sur :
puis leur dissociation contrôlée par l’asymétrie nucléocytoplasmique du cycle Ran.
Gradient Ran comme moteur directionnel
1. Ran est une petite GTPase (~25 kDa) suffisamment petite pour diffuser librement à travers le NPC.
Le gradient nucléocytoplasmique de Ran n’est pas un moteur mécanique, mais un système de réaction-diffusion responsable de la directionnalité du transport actif.
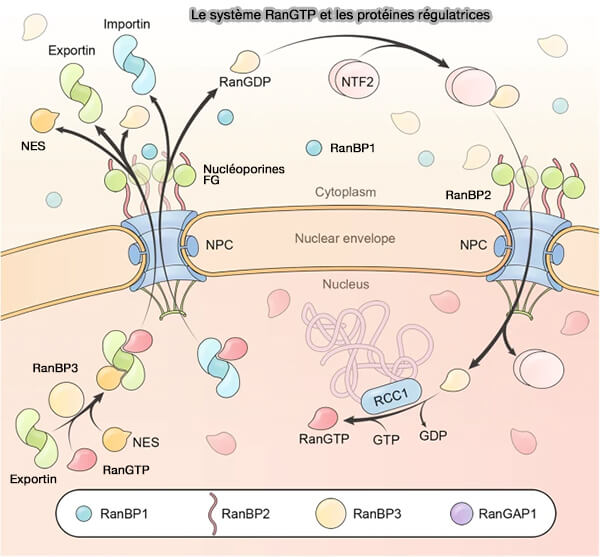
Le système Ran-GTP et ses protéines régulatrices
(Figure : vetopsy.fr d'après Heald et Khodjakov)
2. Ran est présente sous deux formes.
a. Dans le noyau, Ran-GTP est enrichi car RCC1 (RanGEF), liée à la chromatine, catalyse l’échange GDP ➞ GTP, i.e. toute molécule de Ran-GDP qui entre dans le noyau est rapidement convertie en Ran-GTP.
b. Dans le cytoplasme, après exportation de Ran-GTP, Ran-GDP est prédominant en raison de la présence de :
RanBP1 qui se lie spécifiquement à Ran-GTP et facilite son interaction avec RanGAP,
Remarque : RanBP3 se lie aussi à Ran-GTP, mais uniquement pour stabiliser le complexe exportine–Ran-GTP–cargo et moduler l’affinité de l'exportine (CRM1) pour ses substrats.
Cette asymétrie biochimique, maintenue par la séparation spatiale entre RCC1 nucléaire et RanGAP cytoplasmique, crée un environnement nucléaire favorisant l’assemblage ou la dissociation spécifique des complexes de transport, selon qu’il s’agisse d’import ou d’export.
c. Bien que Ran-GDP soit suffisamment petit pour diffuser passivement à travers le NPC, son recyclage efficace vers le noyau repose principalement sur son association à NTF2, qui reconnaît spécifiquement la forme GDP-liée de Ran et interagit avec les nucléoporines riches en motifs FG.
Ce transporteur assure une réimportation rapide et contrôlée de Ran-GDP, où il est ensuite converti en Ran-GTP par RCC1 liée à la chromatine, condition indispensable au maintien du gradient nucléocytoplasmique de Ran-GTP.
Remarque : le gradient de Ran ne se limite pas au transport nucléocytoplasmique.
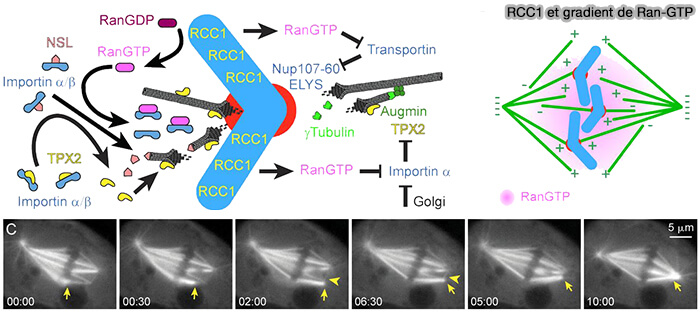
RCC1 et gradient Ran-GTP
(Figure : vetopsy.fr d'après Heald et Khodjakov)
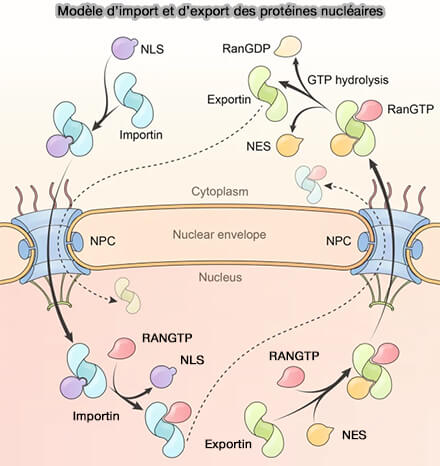
Import et export nucléaires
Le transport nucléocytoplasmique repose sur deux flux directionnels complémentaires assurés par des récepteurs distincts mais fondés sur une logique mécanistique commune.
Modèle d’import et d’export des protéines nucléaires
(Figure : vetopsy.fr d'après Yang et coll)
1. L’import nucléaire correspond au transfert des protéines synthétisées dans le cytoplasme vers le noyau.
2. L’export nucléaire assure la sortie des protéines et complexes ribonucléoprotéiques du noyau vers le cytoplasme.
Il repose sur la reconnaissance de signaux d’export nucléaire (NES) par des karyophérines de type exportine, permettant la formation d’un complexe cargo–récepteur stabilisé par Ran-GTP dans le compartiment nucléaire.
3. Ces deux processus partagent :
l’utilisation de récepteurs appartenant à la famille des karyophérines,
Le tableau suivant met en parallèle les principales étapes et déterminants moléculaires de l’import et de l’export nucléaires, afin d’illustrer leur symétrie structurale et leurs différences mécanistiques fondamentales.