L’import nucléaire est un transport directionnel dépendant du gradient Ran-GTP, assurant l’acheminement sélectif des protéines cytoplasmiques vers le noyau via les karyophérines.
L’import nucléaire constitue le volet entrant du transport nucléocytoplasmique et assure le transfert directionnel des protéines synthétisées dans le cytoplasme vers le compartiment nucléaire.
L’import nucléaire débute par la reconnaissance spécifique des protéines destinées au noyau grâce à des séquences de localisation nucléaire appelées NLS (Nuclear Localization Signal).
a. Les NLS monopartites, constitués d’un seul cluster continu de résidus basiques Lys (K)/Arg (R), s'ancrent principalement dans le site majeur de l'importine α comme celui de :
b. Les NLS bipartites engagent conjointement les sites mineur et majeur via deux clusters basiques séparés par une séquence, assurant un ancrage multivalent du cargo et favorisant le recrutement ultérieur de l’importine β via le domaine IBB de l’importine α comme celui de :
de la nucléoplasmine, protéine nucléaire des ovocytes de Xenopus, KRPAATKKAGQAKKKK,
c. Ces signaux de localisation nucléaire (NLS) dits classiques (cNLS) sont reconnus par la voie d’import dépendante des importines α/β, qui constitue la principale voie d’import nucléaire des protéines solubles.
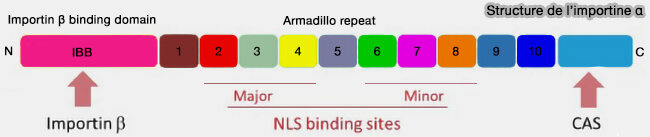
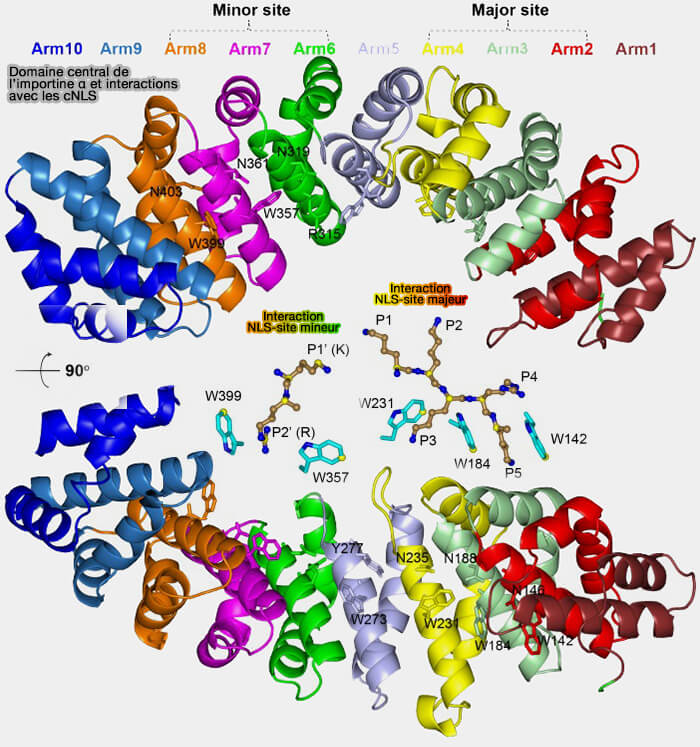
La surface concave formée par les répétitions ARM de l’importine α, qui établit des interactions électrostatiques spécifiques avec les résidus chargés positivement du cargo ( liste des NLS).
a. Les PY-NLS sont caractérisés par 20 à 30 acides aminés qui assument une structure désordonnée, constituée de motifs R/K/H(X)2-5PY C-terminaux avec deux sous-classes selon leurs motifs hydrophobes ou basiques N-terminaux :
hPY-NLS contient des motifs φG/A/Sφφ (où φ est un résidu hydrophobe),
bPY-NLS est enrichi en résidus basiques.
b. D’autres ncNLS existent et sont reconnus par différentes karyophérines β-like selon la nature du cargo.
L’histone H1 est importée via une région C-terminale intrinsèquement désordonnée et fortement enrichie en résidus basiques, reconnue directement par l’importine-7 (IPO7), illustrant une voie d’import indépendante de l’importine α.e.
3. Le masquage ou l’exposition conditionnelle du NLS permet un contrôle dynamique de l’import nucléaire et peuvent être modulés par :
des changements conformationnels, par exemple lors de la dimérisation de certains facteurs de transcription, qui peut exposer un NLS auparavant partiellement enfoui dans la structure tertiaire,
des interactions protéiques, comme pour NF-κB dont le NLS est masqué dans les cellules au repos par son interaction avec IκB ; la phosphorylation puis la dégradation d’IκB exposent le NLS, permettant sa reconnaissance par l’importine α/β et l’import nucléaire,
des modifications post-traductionnelles, comme la phosphorylation de STAT1 suivie de sa dimérisation après stimulation par des cytokines, ce qui stabilise un NLS fonctionnel reconnu par l’importine α5, ou encore certaines modifications post-traductionnelles de p53 qui modulent l’accessibilité de ses signaux nucléaires.
Importines
Les importines sont des protéines de transport spécialisées dans la reconnaissance des signaux de localisation nucléaire (NLS) et dans la traversée du pore nucléaire.
On distingue deux catégories fonctionnelles :
les karyophérines β-like, qui constituent la famille structurale des transporteurs nucléaireset comprennent à la fois des importines et des exportines,
les importines α, adaptateurs spécialisés dans la reconnaissance des NLS classiques.
Remarque : les karyophérines regroupent l’ensemble des récepteurs de transport nucléocytoplasmique dépendants du Ran-GTP.
Karyophérines β-like
Les karyophérines β-like assurent la translocation des complexes cargo–récepteur à travers le pore nucléaire et intègrent cette étape au contrôle directionnel imposé par le gradient nucléocytoplasmique de Ran.
Les exportines appartiennent à la famille structurale des karyophérines β-like. mais elles assurent l’export nucléaire en formant, dans le noyau, un complexe cargo-exportine-Ran-GTP qui est ensuite dissocié dans le cytoplasme après hydrolyse du GTP.
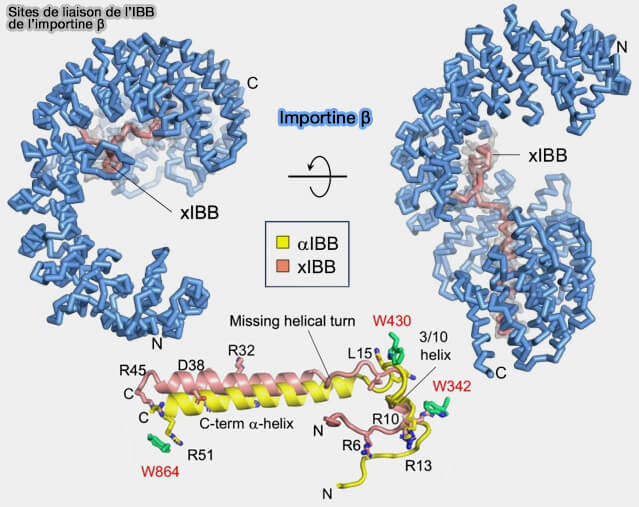
a. Ces domaines IBB (Importin-β Binding) correspondent à des segments N-terminaux riches en résidus basiques capables d’adopter une conformation hélicoïdale lors de leur interaction avec le solénoïde HEAT de l’importine β1.
b. L’αIBB appartient à l’importine α et sert d’interface adaptatrice.
Il permet l’assemblage du complexe trimérique cargo–importine α–importine β en s’insérant dans la cavité interne formée par les répétitions HEAT de β.
L’xIBB inclut souvent des résidus supplémentaires adjacents stabilisant l’interface dans la structure cristallographique.
c. Le sIBB appartient à Snurportin-1 et permet un recrutement direct d’importine β sans intermédiaire α.
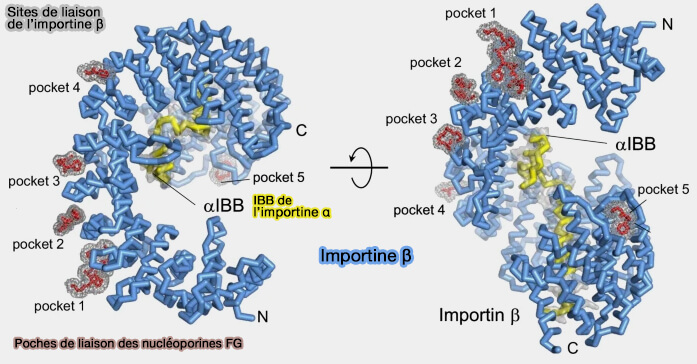
Sites de liaison des FG à l’importine β
(Figure : vetopsy.fr d'après Mitrousis et coll)
Il ne s’agit pas d’un site unique mais de multiples cavités de faible affinité individuelle, dont la coopération multivalente assure une progression dynamique du complexe cargo–récepteur à travers le réseau désordonné des FG-Nup.
Les cinq poches sont situées sur les ARM4/5/6, Arm6/7, ARM7/8, ARM9/10 et ARM16/17.
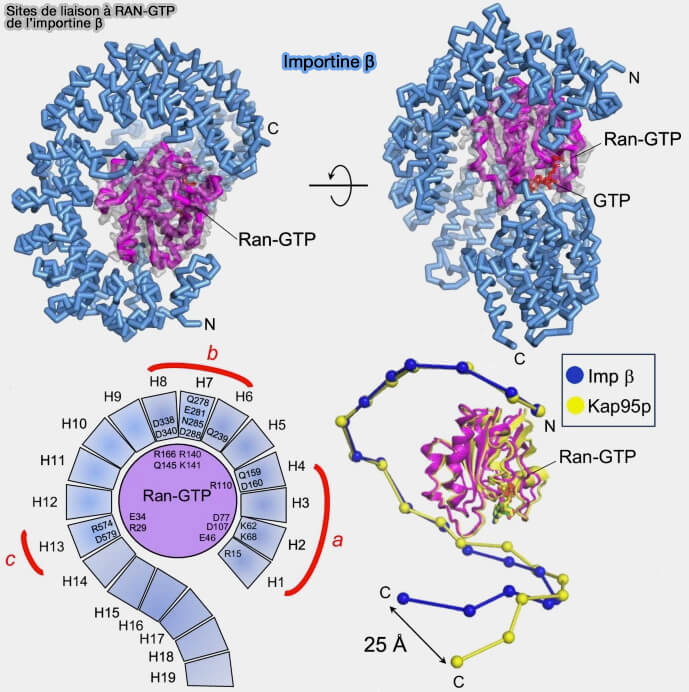
b. Dans le noyau, Ran-GTP se fixe sur la surface concave de l’importine β1 et induit une réorganisation allostérique du solénoïde HEAT.
Cette liaison modifie la géométrie des surfaces concave et convexe impliquées dans la reconnaissance du domaine IBB et des répétitions FG, diminuant leur affinité et favorisant la dissociation du complexe d’import dans le compartiment nucléaire.
Cette modulation conformationnelle constitue la base structurale de la directionnalité du transport nucléocytoplasmique.
Liaison de l’importine β avec RAN-GTP
(Figure : vetopsy.fr d'après Mitrousis et coll)
Les karyophérines β-like assurent la translocation des complexes cargo–récepteur à travers le pore nucléaire et intègrent cette étape au contrôle directionnel imposé par le gradient nucléocytoplasmique de Ran.
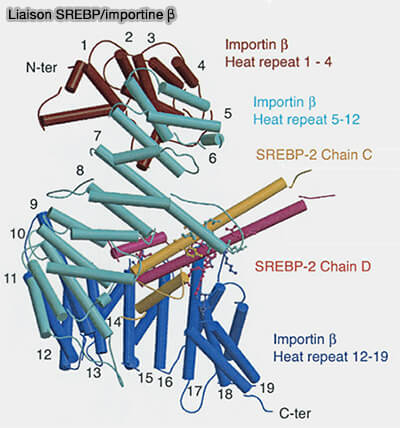
Liaison SREBP/importine β
(Figure : vetopsy.fr d'après Lee et coll)
1. L’importine β1 (KPNB1) constitue le transporteur canonique de la voie dépendante de l’importine α, transportant notamment des facteurs de transcription tels que p53 ou F-κB, ainsi que des histones et des polymérases nucléaires.
3. D’autres karyophérines β-like interviennent dans des voies spécialisées d’import adaptées à certains cargos.
L’importine-7 (IPO7) est capable d’agir comme transporteur direct pour certains cargos, notamment l’histone H1, dont l’import nucléaire est essentiel à l’organisation de la chromatine, et peut, selon la nature du substrat et le contexte cellulaire, coopérer avec d’autres karyophérines β-like afin d’assurer un transport efficace.
L’importine-5 (IPO5) est impliquée dans l’import de l’actine nucléaire et de certains facteurs liés à la dynamique chromatinienne.
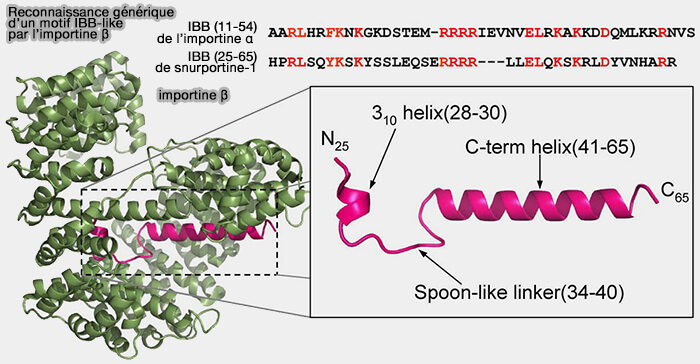
Le " spoon-like linker ", en forme de cuillère, correspond à une boucle flexible qui confère une plasticité conformationnelle permettant l’ajustement du peptide lors de son insertion dans la cavité formée par les répétitions HEAT de l’importine β.
La " 3₁₀-hélice " est une hélice courte et plus resserrée qu’une hélice α classique, qui rigidifie localement le domaine IBB et positionne avec précision les résidus impliqués dans l’interaction avec l’importine β.
Remarque : le domaine IBB, en l’absence d’importine β, peut occuper le site majeur de liaison du NLS sur l’importine α, stabilisant ainsi une conformation auto-inhibée.
Reconnaissance générique d’un motif IBB-like par l’importine β
(Figure : vetopsy.fr d'après Mitrousis et coll)
3. L’importine α ne traverse pas seule le pore nucléaire et dépend de l’importine β1 (KPNB1) pour assurer la translocation du complexe cargo–récepteur à travers le NPC.
Mécanisme de l'import
Arrimage cytoplasmique du complexe d’import
Cette étape correspond à une phase de " docking ", i.e. arrimage, qui positionne le complexe importine–cargo en face de l’axe du pore avant son engagement dans le canal central.
Dans le cytoplasme, les protéines destinées au noyau sont d’abord identifiées par la présence d’un signal de localisation nucléaire (NLS), généralement riche en résidus basiques exposés à la surface du cargo.
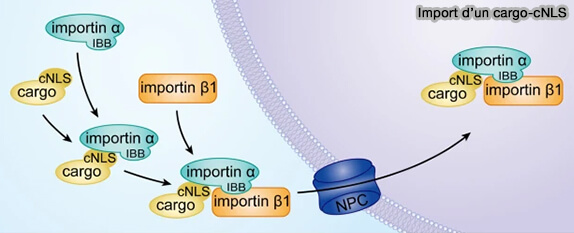
a. Dans le modèle canonique, le NLS est reconnu par l’importine α, dont les répétitions ARM forment une surface concave adaptée à la fixation des séquences basiques.
L’importine β se lie alors au domaine IBB (Importin-β Binding domain) situé à l’extrémité N-terminale de l’importine α, formant un complexe trimérique cargo–α–β compétent pour la translocation.
Import nucléocytoplasmque d'un cargo-cNLS
(Figure : vetopsy.fr d'après Lu et coll)
b. Dans des voies non canoniques, certaines karyophérines β-like reconnaissent directement le cargo sans adaptateur α, en se liant à des signaux structuraux spécifiques tels que les PY-NLS.
Les complexes importine–cargo diffusent librement dans le cytoplasme et leur rencontre avec le NPC dépend à la fois de la diffusion et de la concentration locale des récepteurs.
Les filaments cytoplasmiques constituent une zone périphérique d’interface entre le cytosol et le pore, augmentant la probabilité de capture des complexes d’import.
Grâce à la présence de nucléoporines riches en répétitions FG, ils permettent des interactions transitoires initiales avec les karyophérines, stabilisant temporairement le complexe à la surface du pore avant son engagement dans le canal central.
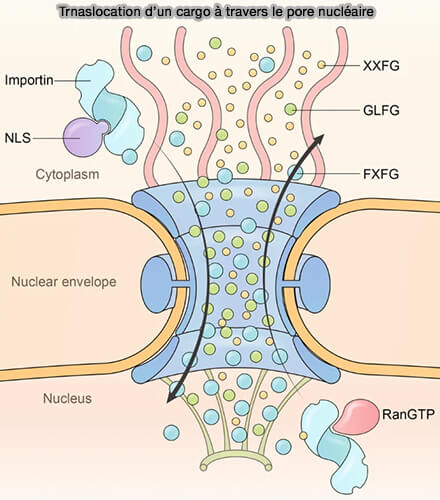
Translocation dans le canal central
Une fois arrimé à la face cytoplasmique du pore, le complexe importine–cargo s’engage dans le canal central et progresse à travers le réseau dense de nucléoporines riches en répétitions FG (phénylalanine-glycine).
Translocation d'un cargo à travers le pore nucléaire
(Figure : vetopsy.fr d'après Yang et coll)
La translocation correspond à une diffusion facilitée à l’intérieur de ce réseau sélectif, dont la perméabilité dépend des interactions spécifiques établies entre les répétitions FG et la surface des karyophérines β-like.
La translocation ne requiert pas d’ATP directement au niveau du pore.
Les protéines volumineuses sans interaction spécifique sont ainsi exclues, tandis que les complexes liés aux karyophérines peuvent progresser.
a. La progression du complexe importine–cargo repose sur des interactions multivalentes et transitoires entre l’importine β-like et les motifs FG.
Les résidus phénylalanine des répétitions FG s’insèrent temporairement dans des poches hydrophobes réparties le long de la surface de l’importine β.
Ces interactions sont de faible affinité individuellement, mais nombreuses et répétées, permettant une succession rapide d’associations et de dissociations.
b. Le transporteur ne se fixe donc pas de manière stable à un site unique, mais établit une série de contacts courts avec différents motifs FG, ce qui permet une progression par " relais " au sein du réseau.
Dissociation nucléaire du complexe d’import
Dans le noyau, Ran sous forme liée au GTP se fixe spécifiquement à l’importine β.
Cette liaison induit un changement conformationnel de l’importine β qui réduit son affinité pour le complexe importine α–cargo.
Cette dissociation nucléaire ne dépend pas de l’hydrolyse du GTP.
1. La conséquence immédiate est la dissociation du complexe trimérique cargo–importine α–importine β en deux complexes :
un complexe importine β–Ran-GTP, stable dans l’environnement nucléaire enrichi en Ran-GTP,
un complexe transitoire importine α-cargo.
2. Nup50, nucléoporine du panier nucléaire, se fixe sur le site majeur de liaison du NLS au sein du sillon ARM de l’importine α, entre en compétition directe avec le cargo et favorise sa libération.
Après libération du cargo, l’importine α devient disponible pour l’export.
Le domaine IBB peut réoccuper transitoirement le site majeur, stabilisant une conformation auto-inhibée.
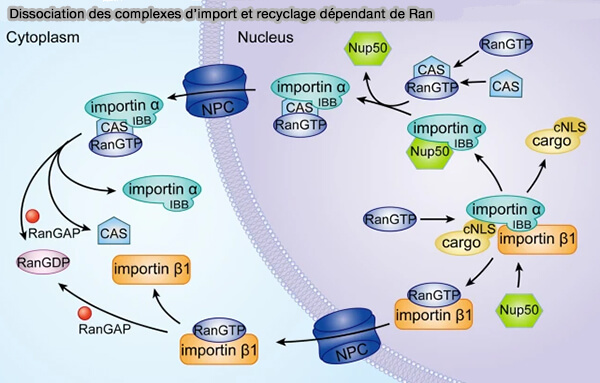
Dissociation des complexes d’import et recyclage dépendant de Ran
1. Le complexe importine β–Ran-GTP, formé dans le noyau après dissociation du cargo, traverse le pore nucléaire vers le cytoplasme par diffusion facilitée, grâce aux interactions répétées entre l’importine β et les nucléoporines riches en motifs FG.
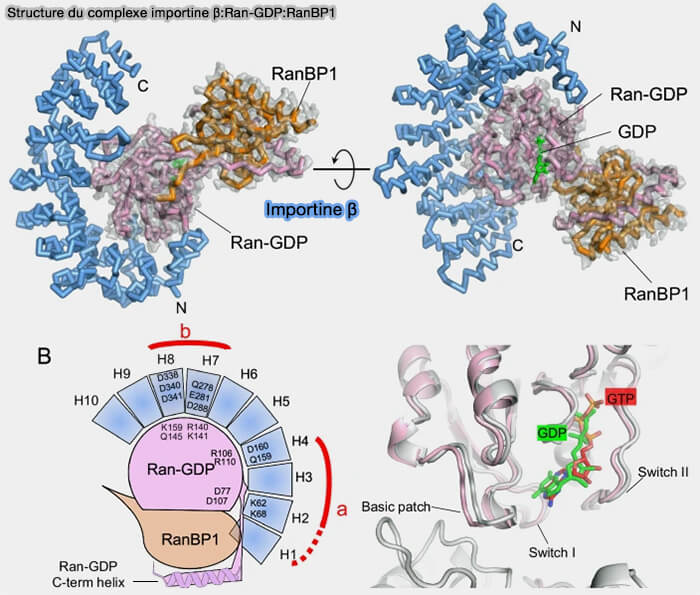
Structure du complexe importine β:Ran-GDP:RanBP1
(Figure : vetopsy.fr d'après Mitrousis et coll)
L’hydrolyse convertit Ran-GTP en Ran-GDP et induit un changement conformationnel qui diminue son affinité pour l’importine β, entraînant la dissociation du complexe.
L’importine β est alors libérée sous forme libre et peut engager un nouveau cycle d’import.
b. Ran-GDP, incapable de diffuser efficacement seul à travers le pore, est pris en charge par le transporteur spécifique NTF2 (Nuclear Transport Factor 2) et retraverse le pore nucléaire dans le sens cytoplasme ➞ noyau pour être transformé en Ran-GTP ( translocation nucléaire de Ran-GDP).
Dissociation des complexes d’import et recyclage dépendant de Ran
(Figure : vetopsy.fr d'après Lu et coll)
2. Après dissociation du cargo dans le noyau, l'importine α est transitoirement associée à Nup50 sous forme de complexe importine α–Nup50 et doit être recyclée.
a. Dans le noyau, CAS (Cellular Apoptosis Susceptibility protein), une importine spécifique qui appartient à la famille des karyophérines β-like, se lie à Ran-GTP.
b. Le complexe CAS–Ran-GTP reconnaît principalement la région C-terminale formée par les répétitions ARM de l’importine α, ce qui entraîne :
la dissociation de Nup50,
la formation du complexe trimérique stable importine α–CAS–Ran-GTP.
c. Ce complexe est exporté vers le cytoplasme par diffusion facilitée à travers le pore nucléaire.
d. Dans le cytoplasme, l’hydrolyse du GTP lié à Ran entraîne une réorganisation conformationnelle de CAS et la dissociation du complexe importine α–CAS–Ran-GTP.
L’importine α et CAS sont ainsi libérées et disponibles pour un nouveau cycle de transport, tandis que Ran-GDP est recyclé vers le noyau via NTF2.
Remarque : une synthèse comparative présente côte à côte les étapes clés et les déterminants moléculaires de l’import et de l’export nucléaires, afin de mettre en évidence leur organisation symétrique ainsi que leurs divergences mécanistiques essentielles.




 liste des NLS).
liste des NLS).