


Noyau
Enveloppe nucléaire : architecture et composition
- Biologie cellulaire et moléculaire
- Constituants de la cellule
- Matrice extracellulaire
- Reproduction cellulaire
- Biochimie
- Transport membranaire
- Moteurs moléculaires

L’enveloppe nucléaire est une double membrane spécialisée qui sépare nucléoplasme et cytoplasme, organise la lamina et la chromatine périphérique et assure la transmission des forces mécaniques via le complexe LINC.
Vue d'ensemble de l'enveloppe nucléaire
L’enveloppe nucléaire constitue une structure hautement spécialisée qui assure la délimitation, la protection et l’intégration mécanique du noyau au sein de la cellule eucaryote.
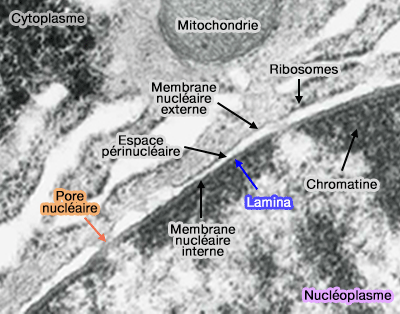
(Figure : vetopsy.fr)
1. L’enveloppe nucléaire est formée par (The Nuclear Envelope 2010) :
- une membrane nucléaire interne,
- un espace périnucléaire,
- une membrane nucléaire externe,
- des complexes de pores nucléaires assurant le transport nucléocytoplasmique.
2. La membrane nucléaire externe et la membrane nucléaire interne sont toutes deux des bicouches lipidiques classiques, organisées selon l’architecture générale des membranes biologiques ( structure de la membrane plasmique).
structure de la membrane plasmique).
La particularité de l'enveloppe nucléaire réside dans sa spécialisation protéique et son organisation mécanique et de son organisation globale, i.e. double membrane séparée par un espace périnucléaire, présence des pores nucléaires, interaction structurale avec la lamina et la chromatine périphérique (Mechanics of nuclear membranes 2019).
2. Elle exerce de multiples fonctions structurales et organisationnelles essentielles à la stabilité et au fonctionnement du compartiment nucléaire.
- Elle articule membranes, lamina nucléaire et chromatine périphérique au sein d’un système structural cohérent.
- Elle relie le noyau au cytosquelette et au réticulum endoplasmique (RE), assurant son insertion dans l’architecture cellulaire globale.
- Elle maintient un compartiment nucléaire fonctionnel tout en permettant une plasticité contrôlée au cours du cycle cellulaire.
- Elle participe à l’organisation tridimensionnelle du génome en interphase et à sa reconfiguration en mitose.
3. L’enveloppe nucléaire apparaît ainsi comme une interface structurale dynamique reliant architecture membranaire, organisation chromatinienne et mécanique cellulaire.

Vous pouvez lire : The nuclear envelope and nuclear pore complexes in neurodegenerative diseases (2025) et Tuning between Nuclear Organization and Functionality in Health and Disease (2023).
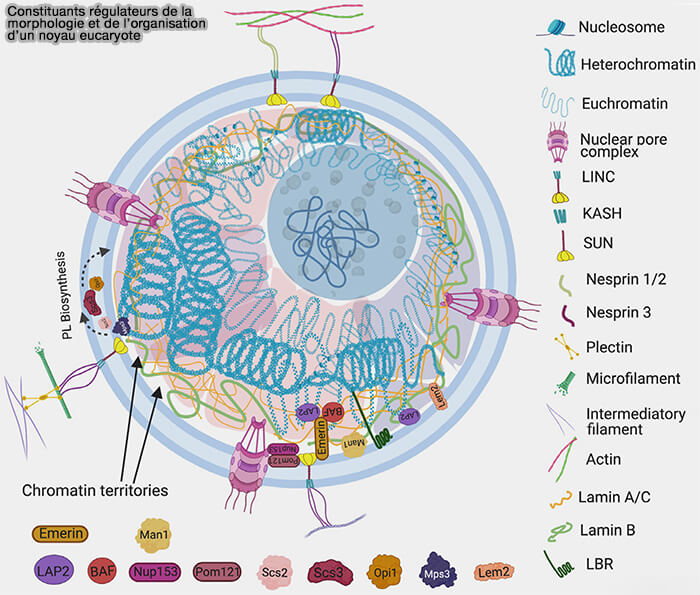
(Figure : vetopsy.fr d'après Manda et coll)
Membrane nucléaire interne
La membrane nucléaire interne constitue une membrane spécialisée, distincte de la membrane externe, enrichie en protéines spécifiques assurant l’organisation structurale du noyau.
Organisation structurale
1. Comme son nom l'indique, elle constitue le feuillet membranaire interne de l’enveloppe nucléaire.
a. Elle est orientée vers le nucléoplasme, ce qui la distingue topologiquement et fonctionnellement de la membrane nucléaire externe.
- Elle est séparée de la membrane externe par l’espace périnucléaire et présente une composition protéique distincte malgré la continuité membranaire avec le réticulum endoplasmique (RE).
- Elle sert de support d’ancrage à la lamina nucléaire, via des protéines transmembranaires interagissant avec les lamines A/C et B.
- Elle établit des connexions directes ou indirectes avec la chromatine périphérique, participant à la stabilisation des LAD (Lamina-Associated Domains) et à l’organisation tridimensionnelle du génome.
b. Elle constitue également une plateforme d’intégration de signaux mécaniques et régulateurs, en interface avec la lamina et le complexe LINC.
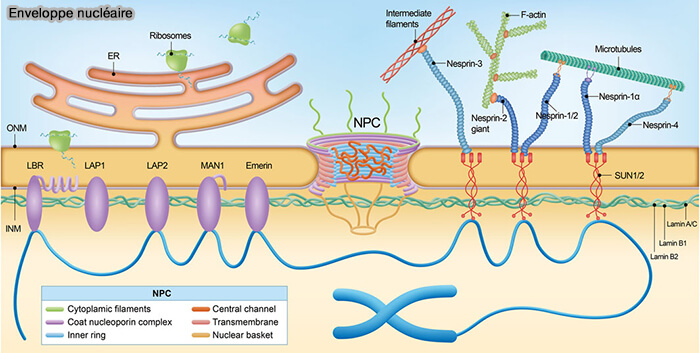
(Figure : vetopsy.fr d'après Wu et coll)
2. La membrane nucléaire interne est caractérisée par un ensemble de protéines transmembranaires qui assurent l’ancrage de la lamina et l’organisation de la chromatine périphérique (protéines de la membrane nucléaire interne associées aux lamines).
a. Les protéines à domaine LEM (LAP2β, émerine, MAN1, LEMD2) établissent un couplage structurant entre lamina et chromatine, notamment via leur interaction avec BAF.
LAP1 et certaines isoformes de LAP2 participent à la stabilisation du réseau laminaire contre la membrane interne.
b. LBR (Lamin B Receptor) interagit avec les lamines B et des composants de l’hétérochromatine, contribuant à l’ancrage des LAD à la périphérie nucléaire.
Remarque : d’autres protéines associées à la lamina modulent la signalisation nucléaire, la mécanique et la réponse aux contraintes cellulaires (partenaires régulateurs et fonctionnels de la lamina).
Protéines de la membrane nucléaire interne
La membrane nucléaire interne constitue un domaine membranaire hautement spécialisé caractérisé par la présence de protéines spécifiques assurant :
- l’ancrage de la lamina nucléaire,
- l’organisation de la chromatine périphérique,
- l’organisation structurale du noyau,
Contrairement à la membrane nucléaire externe, sa composition protéique est fortement différenciée et sa mobilité protéique est restreinte par ses interactions avec les structures nucléaires internes.
Vous pouvez lire : The Diverse Cellular Functions of Inner Nuclear Membrane Proteins (2021).
1. Des protéines assurent l’ancrage direct de la membrane nucléaire interne à la lamina nucléaire (protéines associées aux lamines).
- Ces protéines interagissent avec les lamines A/C et B et contribuent à la stabilité mécanique du noyau, au maintien de sa forme et à l’organisation de l’architecture nucléaire.
- On peut citer notamment LAP1 et des protéines à domaine LEM, comme l’émerine et LAP2β.
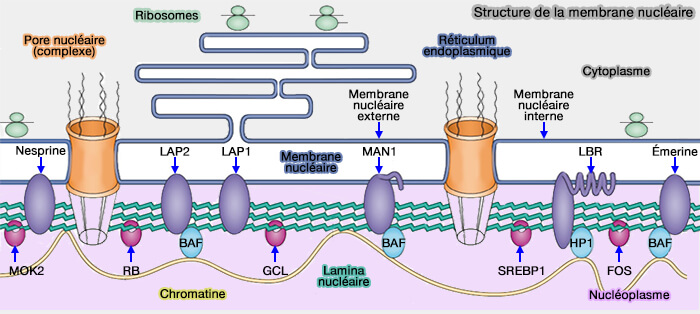
(Figure : vetopsy.fr d'après H. D. M Coutinho)
{kind=link}
3. La membrane nucléaire interne contient également des protéines capables de se lier directement ou indirectement à la chromatine et d’assurer l’ancrage périphérique de régions chromosomiques spécifiques :
- soit par liaison aux lamines, comme LBR (Lamin B receptor), récepteur de la lamine B impliqué dans l’ancrage de l’hétérochromatine périphérique,
- soit via des facteurs adaptateurs tels que BAF dans le cas des protéines à domaine LEM.
Ces interactions contribuent à l’organisation de l’hétérochromatine et participent à la structuration tridimensionnelle du génome ainsi qu’à la distribution spatiale des domaines transcriptionnellement actifs ou réprimés.
4. La membrane nucléaire interne contient les protéines SUN, caractérisées par un domaine nucléoplasmique associé à la lamina et un domaine luminal localisé dans l’espace périnucléaire ( complexe LINC).
- Ces protéines assurent le couplage structural entre membrane nucléaire interne et membrane nucléaire externe par interaction avec des partenaires transmembranaires situés dans la membrane externe.
- Elles permettent ainsi la transmission des contraintes mécaniques à travers l’enveloppe nucléaire et participent au positionnement du noyau dans la cellule.
Fonctions et dynamique de la membrane nucléaire interne
1. La membrane nucléaire interne assure un rôle architectural central dans l’organisation du noyau en interphase.
- Elle stabilise le réseau laminaire périphérique et contribue à la rigidité de l’enveloppe nucléaire.
- Elle participe à l’ancrage sélectif de domaines chromatiniens périphériques, structurant l’interface entre hétérochromatine et lamina.
Par ces interactions, elle contribue indirectement au maintien des LAD et à la stabilisation des territoires chromosomiques.
Elle participe au couplage mécanique du noyau en interaction avec le complexe LINC, qui relie la lamina au cytosquelette à travers les deux membranes nucléaires.
Remarque : la composition et la distribution des protéines de la membrane interne peuvent varier selon le type cellulaire et l’état de différenciation.
Des altérations de ces protéines affectent la stabilité mécanique du noyau et sont impliquées dans diverses pathologies, notamment les laminopathies.
2. La membrane nucléaire interne est soumise à une régulation dynamique étroitement couplée au cycle cellulaire et à l’état fonctionnel de la cellule.
- Au début de la prométaphase, la phosphorylation coordonnée des lamines et des protéines transmembranaires diminue leurs interactions réciproques, favorisant la dissociation de l’interface lamina-membrane-chromatine (rupture de l’enveloppe nucléaire).
- En sortie de mitose, leur déphosphorylation restaure progressivement l’ancrage laminaire et la réassociation des domaines chromatiniens périphériques (reformation de l’enveloppe nucléaire).
Espace périnucléaire
1. L’espace périnucléaire correspond au compartiment situé entre les membranes nucléaires interne et externe et constitue une continuité structurale avec la lumière du réticulum endoplasmique (RE).
- Il présente une largeur généralement comprise entre 20 et 40 nm, assurant une unité topologique du système endomembranaire.
- Il abrite les domaines luminales des protéines transmembranaires des deux membranes.
- Il contient un milieu luminal continu avec celui du réticulum endoplasmique, incluant des ions, des protéines chaperonnes et des enzymes résidentes du RE (The perinuclear region concentrates disordered proteins with predicted phase separation distributed in a 3D network of cytoskeletal filaments and organelles 2022).
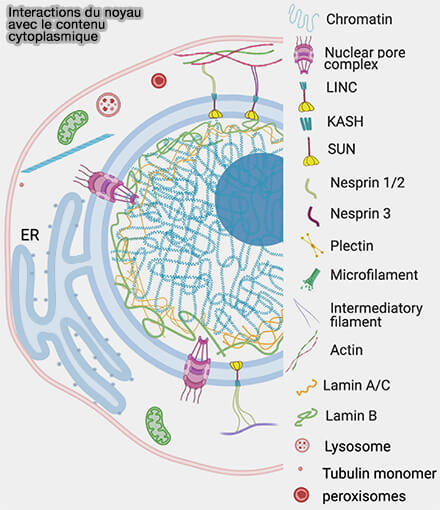
(Figure : vetopsy.fr d'après Manda et coll)
Remarque : les mécanismes assurant le maintien de cet espacement restent encore débattus et il ne semble pas que les protéines SUN jouent un rôle déterminant dans le contrôle direct de cette distance (SUN proteins and nuclear envelope spacing 2015).
2. Il participe à l’organisation mécanique de l’enveloppe nucléaire en servant d’espace de couplage entre les deux membranes, notamment via l’interaction des domaines luminales des protéines SUN avec les domaines KACH des nesprines.
Membrane
nucléaire
externe
La membrane nucléaire externe constitue la composante cytoplasmique de l’enveloppe nucléaire et assure la continuité structurale avec le réticulum endoplasmique rugueux, et partage avec lui une composition lipidique et protéique proche.
Organisation structurale
1. La membrane nucléaire externe représente ainsi une région spécialisée du réseau endomembranaire périphérique du noyau.
- Sa face cytoplasmique peut être associée aux ribosomes, traduisant sa proximité fonctionnelle avec le système de synthèse protéique et la production des protéines destinées aux membranes ou au système sécrétoire.
- L’espace périnucléaire correspond à la lumière du réticulum endoplasmique, assurant une continuité luminale entre enveloppe nucléaire et réticulum et garantissant l’unité topologique du système endomembranaire.
2. Contrairement à la membrane nucléaire interne, elle ne porte pas les protéines d’ancrage spécifiques de la lamina ni les complexes d’interaction directe avec la chromatine.
Elle participe à l’organisation architecturale de l’enveloppe nucléaire et au couplage mécanique du noyau avec le cytosquelette, contribuant à l’intégration du noyau dans l’architecture cellulaire globale.
Protéines de la membrane nucléaire externe
La membrane nucléaire externe présente une composition protéique proche de celle du réticulum endoplasmique (RE), mais contient également des protéines spécifiques assurant l’interface entre le noyau et le cytoplasme ainsi que le couplage mécanique avec le cytosquelette.
1. La membrane nucléaire externe partage de nombreuses protéines avec le réticulum endoplasmique, reflétant leur continuité topologique.
Elle contient notamment des composants impliqués dans :
- la synthèse et l’insertion des protéines, comme le translocon Sec61 et ses complexes associés (TRAM, TRAP) qui reflètent l’association de la face cytoplasmique de la membrane nucléaire externe aux ribosomes et sa continuité structurale et fonctionnelle avec le réticulum endoplasmique,
- la biosynthèse lipidique, comme les enzymes de synthèse des phospholipides ( synthèses de la phosphatidylcholine et de la phosphatidyléthanolamine) et les enzymes de synthèse du cholestérol,
- le transport membranaire vers l'appareil de Golgi, via la machinerie COPII, et les protéines associées aux sites de sortie du RE comme TANGO1, ainsi que le trafic rétrograde COPI assurant le recyclage vers le RE et le passage par le compartiment intermédiaire ERGIC.
2. La membrane nucléaire externe contient des protéines transmembranaires impliquées dans le couplage mécanique du noyau avec le cytosquelette ( complexe LINC).
- Parmi celles-ci figurent notamment les nesprines, protéines à domaine KASH, dont le domaine luminal interagit avec les protéines SUN de la membrane nucléaire interne.
- Ces interactions assurent la continuité mécanique entre cytosquelette, enveloppe nucléaire et lamina, permettant la transmission des forces mécaniques, le positionnement du noyau et l’organisation spatiale des structures cellulaires.
3. Certaines protéines de la membrane nucléaire externe participent à l’organisation structurale et à la dynamique de l’enveloppe nucléaire, notamment lors de sa réorganisation au cours du cycle cellulaire.
Elles contribuent au maintien de la continuité membranaire avec le réticulum endoplasmique et à l’intégrité globale de l’enveloppe nucléaire.
a. Les torsines, AAA+ ATPases localisées dans l’espace périnucléaire, participent au remodelage de l’enveloppe nucléaire en facilitant l’assemblage et la maturation des complexes de pores nucléaires et en contribuant au maintien de l’intégrité de la membrane.
Leur activité dépend des cofacteurs membranaires LAP1 (membrane nucléaire interne) et LULL1 (membrane nucléaire externe), qui stimulent leur activité ATPasique.
b. Les réticulons et les protéines DP1/Yop1 sont des protéines insérées en épingle à cheveux dans la membrane, générant une courbure positive élevée.
Elles stabilisent les régions fortement incurvées du réticulum endoplasmique et participent indirectement à l’architecture et à la continuité géométrique entre RE et enveloppe nucléaire.
Différences entre les membranes
Les membranes du réticulum endoplasmique, de la membrane nucléaire externe et de la membrane nucléaire interne présentent une organisation commune en bicouche lipidique, mais se distinguent par leur continuité topologique, leur spécialisation protéique et leurs fonctions structurales.
Le tableau suivant compare leurs propriétés organisationnelles, leurs interactions et leurs rôles biologiques afin de préciser leur degré de différenciation au sein du système endomembranaire et de l’enveloppe nucléaire.
| Caractéristique | Membrane du réticulum endoplasmique (RE) |
Membrane nucléaire externe |
Membrane nucléaire interne |
|---|---|---|---|
| Position | Réseau endomembranaire cytoplasmique (tubules et citernes) | Face cytoplasmique de l’enveloppe nucléaire | Face nucléoplasmique de l’enveloppe nucléaire |
| Organisation membranaire | Bicouche lipidique simple | Bicouche lipidique simple | Bicouche lipidique simple |
| Organisation globale | Réseau continu dynamique | Partie de la double membrane nucléaire | Partie de la double membrane nucléaire |
| Continuité membranaire | Continue avec membrane nucléaire externe | Continue avec RE et espace périnucléaire | Non continue avec RE (séparée fonctionnellement) |
| Continuité luminale | Lumière du RE | Espace périnucléaire = lumière du RE | Espace périnucléaire du côté luminal |
| Composition lipidique | Composition standard du système endomembranaire | Très proche du RE | Composition spécialisée |
| Spécialisation protéique |
|
|
Protéines spécifiques d’organisation nucléaire |
| Association aux ribosomes | Oui (RE rugueux) | Possible sur face cytoplasmique | Non |
| Interaction avec la chromatine | Aucune | Aucune directe | Oui (organisation chromatine périphérique) |
| Interaction avec la lamina | Non | Non | Oui par ancrage direct |
| Interaction cytosquelette | Limitée | Importante (couplage mécanique du noyau) | Indirecte |
| Mobilité des protéines | Élevée | Relativement élevée | Fortement restreinte |
| Rôle principal |
|
|
Organisation nucléaire et structurale |
| Contraintes mécaniques | Faibles à modérées | Importantes (positionnement nucléaire) | Très élevées (stabilité nucléaire) |
| Fonction structurale nucléaire | Aucune | Support architectural externe | Organisation interne du noyau |
| Spécificité biologique | Organelle métabolique | Extension spécialisée du RE | Domaine membranaire nucléaire spécialisé |
Rôles de l'enveloppe nucléaire
Biologie cellulaire et moléculaireMembrane plasmiqueNoyauEnveloppe nucléairePores nucléairesLaminaCorps nucléairesTransport nucléocytoplasmiqueCytoplasmeMitochondriesSystème endomembranaireRéticulum endoplasmiqueAppareil de GolgiEndosomesLysosomesPeroxysomesProtéasomesCytosqueletteFilaments d'actineFilaments intermédiairesMicrotubulesReproduction cellulaireBiochimieTransport membranaire Moteurs moléculairesVoies de signalisation