Les lamines nucléaires sont des filaments intermédiaires de type V qui forment la lamina nucléaire et structurent l’architecture, la mécanique et l’organisation fonctionnelle du noyau.
Localisation des lamines
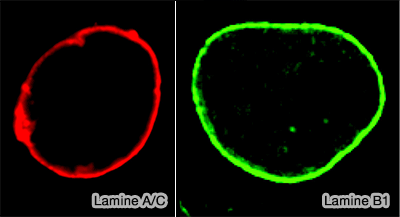
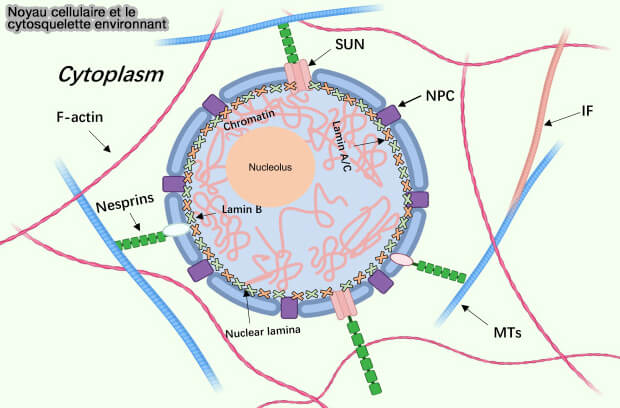
(Figure : vetopsy.fr d'après Dittmer)la lamina nucléaire, armature protéique de 10 à 30 nm de large qui borde l'enveloppe nucléaire sur sa face interne, constituée par l’assemblage des différents types de lamines,
le " voile nucléoplasmique " plus diffus, présent dans le nucléoplasme au cours de l’interphase, principalement associé aux lamines de type A/C, dont une fraction significative n’est pas strictement confinée à la périphérie nucléaire.
1. Les lamines, filaments intermédiaires de classe V, sont classées en deux grands groupes.
a.
Les lamines de type A (lamines A et C) sont uniquement exprimées après la gastrulation (gène LMNA).
Outre les lamines A et C, on identifie des isoformes mineures, telles que la lamine C2 (spécifique du testicule) et la lamine AΔ10 dans certaines cellules somatiques.
Ces réseaux ne sont toutefois pas indépendants et s’interpénètrent partiellement et établissent des interfaces fonctionnelles, contribuant conjointement à l’architecture nucléaire, à l’ancrage de la chromatine et à la compartimentation fonctionnelle du noyau.
3. Bien que les lamines soient des constituants strictement nucléaires à l’état assemblé, leur synthèse a lieu dans le cytoplasme.
Après traduction, les lamines sont importées dans le noyau par le système d’import nucléaire grâce à un signal de localisation nucléaire (NLS) porté par leur domaine C-terminal, avant leur incorporation dans la lamina nucléaire ou le réseau nucléoplasmique.
Structure et polymérisation des lamines
Caractéristiques structurales des lamines nucléaires
1. Les lamines nucléaires présentent des caractéristiques structurales spécifiques qui les distinguent des filaments intermédiaires cytoplasmiques, tout en conservant l’architecture générale des protéines de filaments intermédiaires (Structural basis for lamin assembly at the molecular level 2019).
Structure des lamines A et C
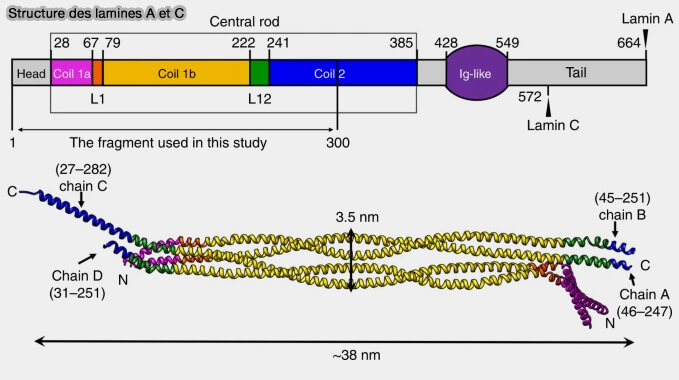
(Figure : vetopsy d'après Ann et coll)
a. Le domaine hélical en forme de tige (rod domain), plus long de 42 acides aminés que celui des filaments intermédiaires cytoplasmiques, est flanqué de part et d'autre d'un site de phosphorylation.
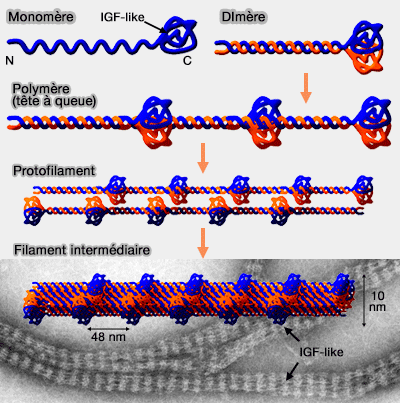
b. Cette polymérisation tête-à-queue permet la formation de polymères linéaires et d’associations de tétramères antiparallèles, au sein desquels le domaine Ig-like globulaire apparaît de manière périodique, avec une répétition d’environ 48 nm.
2. Contrairement aux filaments intermédiaires cytoplasmiques, relativement stables, les filaments intermédiaires nucléaires présentent une dynamique d’assemblage et de désassemblage élevée, en particulier au cours du cycle cellulaire.
La biogenèse et la maturation des lamines reposent sur une succession de modifications post-traductionnelles qui conditionnent leur adressage, leur ancrage membranaire et leur intégration au réseau laminaire.
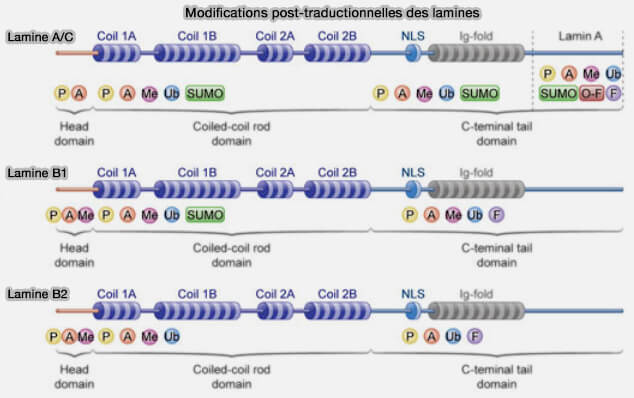
Modifications post-traductionnelles des lamines
(figure : vetopsy.fr d'après Zheng et col)
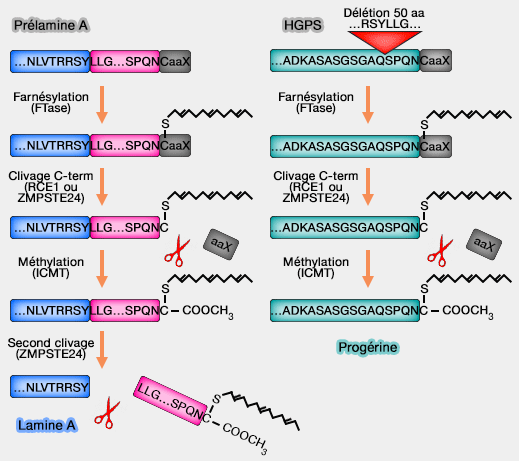
1. La lamine A (664 acides aminés) est synthétisée sous forme de précurseur (prélamine A).
a. La prélamine A (566 + 98 acides aminés) possède une extension C-terminale contenant une boîte CaaX (Cyst, acide aminé aliphatique, X, acide aminé quelconque, mais en général la méthionine).
Ce motif CaaX constitue un signal de modification post-traductionnelle déclenchant l’ajout d’un groupement isoprénylique sur la cystéine terminale, ici un farnésyle (C15), par formation d’une liaison thioéther, étape nécessaire à son association transitoire avec la membrane nucléaire interne.
Remarque : la farnésylation correspond à un type spécifique d’isoprénylation, l’autre forme majeure est la géranylgéranylation, qui implique l’ajout d’un groupement isoprénoïde plus long (C20).
b. La maturation de la lamine A implique successivement :
un clivage protéolytique final éliminant 15 acides aminés, conduisant à la lamine A mature non farnésylée.
c. Le dernier clivage, assuré par la métalloprotéase ZMPSTE24, supprime le segment C-terminal contenant la cystéine farnésylée, ce qui élimine définitivement la boîte CaaX et empêche tout ancrage lipidique permanent de la lamine A mature.
Remarque : la progérine correspond à une forme mutée de la prélamine A, dans laquelle une délétion d’environ 50 acides aminés empêche le clivage final, entraînant le maintien de la farnésylation.
a. La lamine C résulte d’un épissage alternatif du gène LMNA qui introduit un exon terminal spécifique et un codon stop prématuré, supprimant ainsi l’extension C-terminale présente dans la prélamine A.
b. L’absence de motif CaaX exclut toute isoprénylation et tout ancrage lipidique transitoire à la membrane nucléaire interne.
La lamine C s’associe donc exclusivement par interactions protéine–protéine au réseau laminaire et aux partenaires nucléaires, ce qui explique sa distribution plus diffuse dans le nucléoplasme et la formation d’un " voile nucléoplasmique " distinct du réseau périphérique fortement ancré des lamines B.
3. Les lamines B (B1 : 586 acides aminés, B2 : 600) conservent une boîte CaaX sui n’est jamais retirée après maturation, et restent farnésylées.
Ce groupement isoprénylique assure un ancrage lipidique constitutif aux membranes dérivées de l’enveloppe nucléaire, y compris durant la mitose.
La présence ou l’absence d’ancrage lipidique constitue ainsi un déterminant majeur de la compartimentation des lamines, opposant le confinement périphérique stable des lamines B à la distribution plus diffuse et dynamique des lamines A/C dans le nucléoplasme.