L’inhibition des complexes cycline/CDK repose sur des verrous moléculaires, la dégradation des cyclines, le contrôle spatial et des boucles négatives qui assurent la stabilité du cycle cellulaire.
Vue d'ensemble de l'inhibition des complexes cycline/CDK
L’inhibition des complexes cycline/CDK constitue un mécanisme actif et structurant du contrôle du cycle, et non une simple absence d’activation.
1. Un complexe peut être inhibé à plusieurs niveaux :
la possibilité d’arrêt en cas d’anomalie, induction de p21CIP1 par p53 pendant la phase G1 ou la phase S après dommages à l’ADN,
la coordination avec les checkpoints, inhibition de Cdc25 par le voies ATR/Chk1 et ATM/Chk2 pendant la phase S et la phase G2
la stabilité globale du système, extinction irréversible de CDK1 par dégradation de la cycline B via APC/CCdh1 pendant la sortie de mitose et la phase G1 précoce
L’équilibre entre activation et inhibition constitue ainsi la base moléculaire de la fiabilité du cycle cellulaire.
Inhibition par contrôle des cyclines
La cycline constitue elle-même un point majeur de régulation négative, car sa disponibilité conditionne directement l’existence et l’activité du complexe cycline/CDK.
1. L’activité d’un complexe peut être éteinte par l’abaissement du niveau de cycline par plusieurs mécanismes.
a. La dégradation ciblée par les ligases E3 (SCF, APC/C) constitue un mécanisme central d’extinction de l’activité CDK.
La cycline E est ubiquitinée par SCFFbxw7 après phosphorylation, ce qui conduit à l’extinction de l’activité de CDK2 au cours de la phase S.
En quiescence (G0), les cyclines D, E, A et B sont faiblement exprimées, ce qui maintient globalement l’activité CDK à un niveau bas.
c. La stabilité des cyclines peut également dépendre de cofacteurs.
La cycline D1 devient instable en l’absence de signalisation Ras/MAPK, ce qui favorise son export nucléaire puis sa dégradation.
Inversement, la protéine p27KIP1 peut stabiliser certains complexes cycline D/CDK4, et sa dégradation peut donc indirectement contribuer à la déstabilisation de ces complexes.
2. L’activité d’un complexe peut aussi être neutralisée par contrôle spatial.
a. L’exclusion nucléaire constitue un mécanisme de séparation fonctionnelle entre les complexes cycline/CDK et leurs substrats.
Lors de l’entrée en mitose, la phosphorylation de la cycline B1 inactive le NES, ce qui entraîne une accumulation nucléaire brutale du complexe cycline B/CDK1.
b. La séquestration cytoplasmique de régulateurs peut également neutraliser indirectement l’activité des CDK.
La p27KIP1 phosphorylée sur Thr198 est retenue dans le cytoplasme, ce qui diminue son activité inhibitrice sur les complexes cycline/CDK nucléaires.
La Cdc25C phosphorylée par Chk1 se lie aux protéines 14-3-3, est retenue dans le cytoplasme et ne peut plus activer CDK1 par déphosphorylation.
c. La relocalisation de certains complexes vers des compartiments spécifiques permet enfin de restreindre spatialement leur activité.
Ce niveau de contrôle permet une extinction rapide, directionnelle et souvent irréversible de l’activité des CDK, en particulier lors des transitions majeures du cycle, notamment la sortie de mitose et la réinitialisation en phase G1.
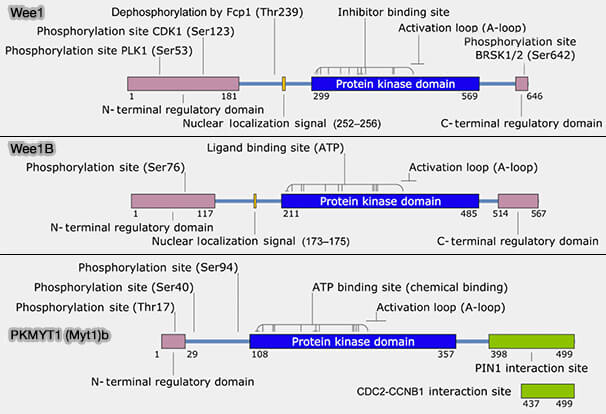
Inhibition par phosphorylation des CDK : Wee1 et Myt1 (PKMYT1)
Certaines phosphorylations bloquent l’activité des complexes cycline/CDK même lorsqu'ils sont correctement assemblés et phosphorylés sur la T-loop par la CAK.
Ce verrou inhibiteur est assuré principalement par deux kinases spécialisées, Wee1 et Myt1.
La kinase Wee1 doit son nom aux mutants " wee " de la levure S. pombe qui entraient prématurément en mitose et produisaient des cellules de très petite taille, i.e. " weel little ”.
Myt1 est l’acronyme de Membrane-associated tyrosine/threonine kinase 1, une kinase apparentée à Wee1, localisée principalement aux membranes, qui participe également à l’inhibition de CDK1.
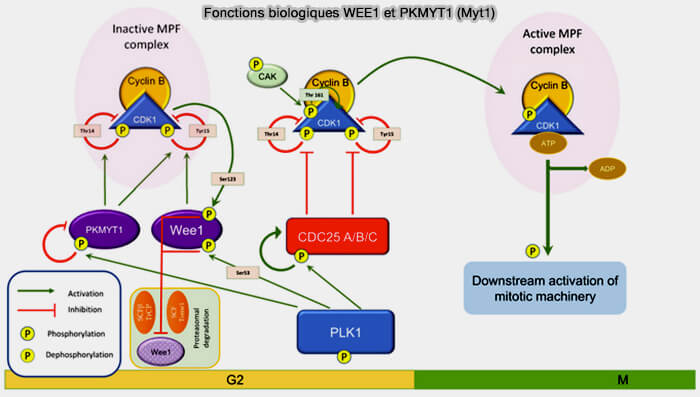
Fonctions biologiques de Wee1 et Myt1 (PKMYT1)
(Figure modifiée : vetopsy.fr d'après di Rorà et coll)
Elle est fortement régulée par phosphorylation dépendante de CDK1 et de Plk1, ce qui contribue à sa désactivation rapide lors de l’entrée en mitose.
Remarque : WEE1B (également appelée WEE2) est une kinase apparentée à WEE1, exprimée spécifiquement dans les ovocytes, et pas dans les cellules somatiques, où elle maintient l’arrêt au stade diplotène de la prophase I en inhibant l’activité du complexe cycline B/CDK1.
b. La CDK demeure alors dans une conformation catalytiquement incompétente, même si la cycline est liée et même si la T-loop est phosphorylée par la CAK.
Fonctions biologiques de Wee1 et Myt1 (PKMYT1)
(Figure modifiée : vetopsy.fr d'après di Rorà et coll)
3. Ce verrou moléculaire constitue un élément central de la chronologie du cycle cellulaire en empêchant :
l’entrée prématurée en mitose avant l’achèvement de la réplication et la préparation structurale des chromosomes,
l’initiation inappropriée de la phase S en l’absence de conditions favorables,
l’engagement de programmes irréversibles tant que l’état cellulaire n’est pas compatible avec la transition suivante.
4. La levée de cette inhibition par Cdc25 représente ainsi un véritable point de bascule fonctionnel, transformant un complexe compétent mais verrouillé, en complexe effectivement actif, par déphosphorylation directe des résidus Thr14 et Tyr15 (Levée d’inhibition etdéclenchement de l’activité CDK).
1. Les checkpoints ne fonctionnent pas comme des voies parallèles autonomes, mais comme des modules de contrôle qui exercent leurs effets principalement par la modulation de l’activité des complexes cycline/CDK.
Au checkpoint S, l’ajustement de la progression de la réplication repose sur une modulation de l’activité de CDK2, associée notamment à l’inhibition transitoire de Cdc25A et à la stabilisation de p27KIP1.
2. Ce niveau d’intégration explique que les CDK ne soient pas de simples enzymes d’exécution du cycle, mais de véritables capteurs et intégrateurs de l’état global de la cellule.
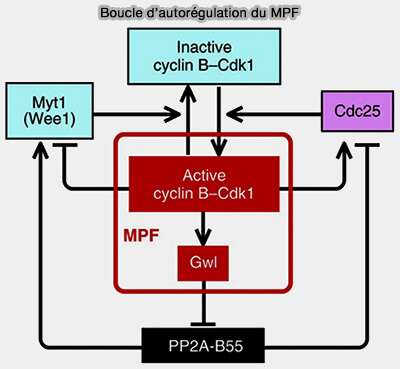
Boucles de rétrocontrôle négatif
Les boucles de rétrocontrôle négatif assurent l’extinction des signaux et la réversibilité des états fonctionnels.
1. Elles reposent principalement sur :
l’activation des systèmes de dégradation des cyclines, comme l’activation de l’APC/C par CDK1 conduisant à la dégradation de la cycline B, qui permet la sortie de mitose,
l’induction de CKI (CDK inhibitors) en réponse aux signaux de stress, comme l’axe p53➞p21CIP1, qui impose un arrêt du cycle et favorise le retour à un état basal en phase G1,
l’élimination programmée d’inhibiteurs lors des transitions, telle que la dégradation de p27KIP1 à l’entrée en phase S, qui contribue à prévenir les activations prolongées ou inappropriées de l’activité CDK.
2. À l’échelle du système, ces boucles négatives assurent la stabilité du cycle cellulaire.
Elles évitent les fluctuations incontrôlées de l’activité CDK, facilitent le retour vers un état basal après une phase active et contribuent à la robustesse globale de la progression du cycle.
Combinées aux boucles positives, elles expliquent que certaines transitions du cycle soient nettes, directionnelles et difficilement réversibles : une fois un seuil franchi, l’état cellulaire bascule de manière coordonnée (par exemple l’entrée en mitose), puis la sortie nécessite l’activation de mécanismes d’extinction spécifiques.




 CKI ou CDK inhibitors),
CKI ou CDK inhibitors),