Les CKI (CDK inhibitors) sont des inhibiteurs endogènes des complexes cycline/CDK qui assurent le freinage du cycle cellulaire en réponse aux signaux de différenciation, de stress ou de dommages de l’ADN.
1. L’inhibition des complexes cycline/CDK constitue un mécanisme actif et structurant du contrôle du cycle, et non une simple absence d’activation.
En outre, un complexe peut être inhibé à d'autres niveaux :
par contrôle de la disponibilité de la cycline, via sa dégradation ou sa non-accumulation (contrôle par les cyclines)
après formation, par blocage catalytique de l’activité kinase (Wee1 et Myt1),
Ils se répartissent en deux grandes familles fonctionnelles :
la famille INK4 (Inhibitor of CDK4), comprenant p15INK4B, p16INK4A, p18INK4C, p19INK4D, qui se lient spécifiquement aux CDK4 et CDK6 et empêchent leur association avec les cyclines D,
La famille INK4 (Inhibitor of CDK4) est strictement spécifique à CDK4 et CDK6 et contrôle le point de restriction R en empêchant leur activation.
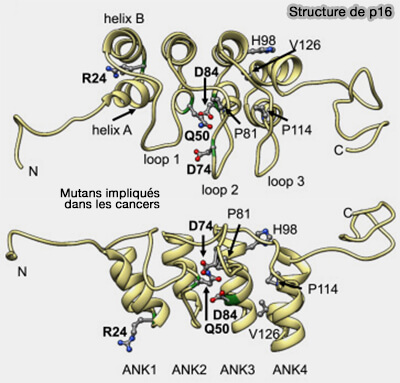
1. Les protéines INK4, p15INK4B, p16INK4A, p18INK4C, p19INK4D, se lient à la CDK monomérique en absence de cycline.
a. Elles sont presque entièrement constituées de répétitions ankyrine (4 à 5 motifs), ce qui en fait des inhibiteurs structurellement rigides, spécialisés dans la déformation de CDK4-6, contrairement aux CIP/KIP qui sont en grande partie désordonnées.
b. Elles sont surexprimées de manière compensatoire dans certaines lésions associées aux HPV (Human Papillomavirus), types de papillomavirus à haut risque capables de favoriser la cancérisation des cellules infectées.
p16 bloque normalement CDK4/6 qui ne phosphorylent plus Rb ➞ Rb reste active ➞ Rb bloque E2F ➞ les divisions sont arrêtées.
E7 désactive directement Rb, donc même si p16 bloque CDK4/6, E2F reste actif et la cellule prolifère.
c. Elles sont fortement exprimées lors de la sénescence cellulaire, ce qui contribue à imposer un arrêt durable du cycle cellulaire et à stabiliser l’état sénescent en empêchant toute ré-entrée en prolifération.
Ce phénomène s’accompagne d’un élargissement de la cellule, d’un aplatissement morphologique et d’une réorganisation profonde de la chromatine, et s’associe au développement d’un phénotype sécrétoire dit SASP (Senescence-Associated Secretory Phenotype), caractérisé par la production de nombreuses molécules pro-inflammatoires pouvant contribuer à des pathologies dégénératives.
Remarque : l’expression de p16INK4A est ainsi largement utilisée comme marqueur cellulaire de ces états biologiques.
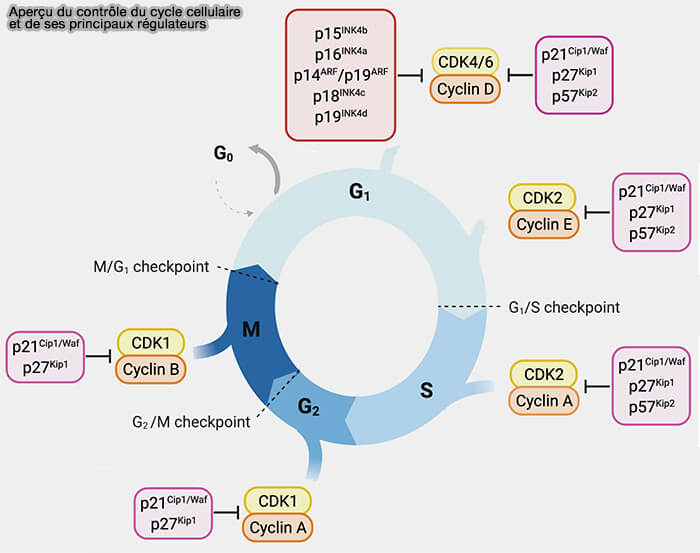
Contrôle du cycle cellulaire et CKI
(Figure modifiée : vetopsy.fr d'après Schirripa et coll)
Famille CIP/KIP : p21, p27, p57
La famille CIP/KIP comprend trois inhibiteurs :
p21CIP1, p27KIP1, p57KIP2.
Contrairement aux INK4, ces protéines ne sont pas spécifiques d’une seule CDK, mais peuvent inhiber plusieurs complexes cycline/CDK.
Structure générale
1. Les protéines CIP/KIP sont majoritairement intrinsèquement désordonnées en solution et adoptent une structure ordonnée uniquement lors de la liaison au complexe cycline/CDK.
b. Les protéines CIP/KIP se lient au complexe déjà assemblé cycline/CDK pour former un complexe ternaire stable cycline/CDK/CKI.
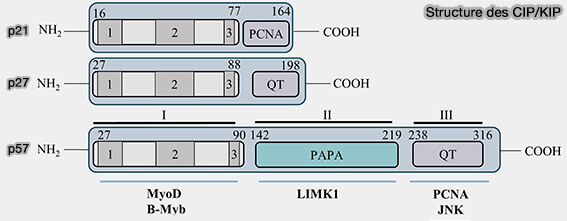
Structure des protéines CIP/KIP
(Figure modifiée : vetopsy.fr d'après Creff et Besson)
2. Leur domaine N-terminal, I sur la figure, constitue un module inhibiteur conservé.
a. Il est organisé en plusieurs sous-régions fonctionnelles :
une région D1 (ou 1), impliquée dans la liaison à la cycline, encore renforcée par la sous-région D1c dans p21, contient ou chevauche un motif court de type RXL (CRS) qui interagit avec la poche de recrutement des substrats de la cycline, renforçant l’ancrage du complexe et contribuant à l’inhibition fonctionnelle.
une région LH (ou 2) qui joue un rôle structural en positionnant correctement D2, le domaine de liaison au CDK,
une région D2 (ou 3), incluant l’hélice 3₁₀, qui pénètre dans la fente catalytique de la CDK, ce qui permet d’obstruer partiellement la poche de liaison à l’ATP, de perturber la géométrie catalytique et d’inhiber
directement la phosphorylation des substrats.
b. Même si la CDK conservait une activité catalytique résiduelle, le complexe cycline/CDK associé à une protéine CIP/KIP ne peut plus recruter correctement les substrats.
3. Leur domaine C-terminal est variable.
p21 contient un motif fonctionnel de liaison à PCNA, facteur central de la réplication de l’ADN, servant de plateforme d’ancrage aux polymérases et aux protéines de réparation, ce qui lui permet d’inhiber directement la réplication de l’ADN indépendamment des CDK.
p27 et p57 possèdent un motif QT conservé, impliqué dans la régulation de leur stabilité, de leur localisation et de certaines interactions protéiques spécifiques, sans rôle direct dans l’inhibition catalytique des CDK.
Remarque : p57 possède une région centrale distincte riche en Pro/Ala chez l’humain, i.e. PAPA qui n’est pas directement impliquée dans l’inhibition des CDK, mais peut engager des interactions spécifiques avec d’autres partenaires.
Fonctions spécifiques de p21, p27 et p57
Bien que partageant un mécanisme d’inhibition commun, p21, p27 et p57 assurent des fonctions biologiques distinctes selon le contexte cellulaire.
p21 assure le lien entre intégrité génomique et contrôle du cycle, en relayant l’activation de p53 vers une inhibition directe des complexes cycline/CDK et, dans certains contextes, de la réplication via PCNA.
p27 agit comme régulateur central de la quiescence (G0) et du point de restriction, en modulant finement le niveau d’activité CDK selon son abondance, sa stabilité et sa localisation.
p57 exerce un rôle spécialisé dans les contextes développementaux, où elle participe au contrôle de la prolifération, de la différenciation et de la maturation tissulaire.




 contrôle par les cyclines)
contrôle par les cyclines)