2. Des mutations de résidus dans les répétitions ANK des protéines sont associées à un grand nombre de maladies humaines : cancer, troubles neurologiques, dysplasies squelettiques…
une boucle (loop ou turn) qui termine la répétition pour se lier au coude β de la répétition suivante, en formant une épingle à cheveux β.
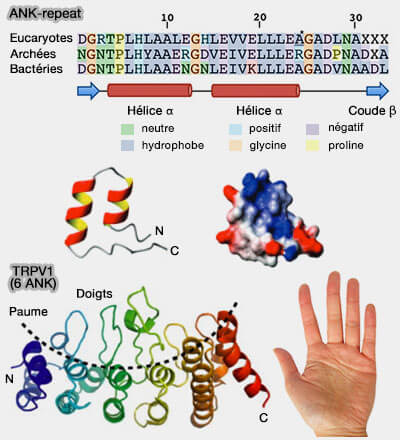
a. Thr-Pro-Leu-His aux positions 6 à 9 forme le coude serré qui initie la première hélice α.
La liaison hydrogène entre le groupe hydroxyle de Thr et le cycle imidazole de His contribue à la stabilité de la répétition motif
et des protéines la contenant.
b. Val/Ile-Val-X (hydrophile)-Leu/Val-Leu-Leu (positions
17 à 22) est la pièce centrale de la deuxième hélice α.
Elle permet de former des liaisons hydrophobes intra- et inter-répétitions pour stabiliser la protéine dans sa globalité.
c. Les résidus conservés de glycine aux positions 13 et 25 terminent les hélices et permettent un pont disulfure pour relier les deux hélices, i.e cette liaison permet à la protéine de se déplier et de se replier.
2. Ces motifs s'empilent les uns sur les autres, en formant des faisceaux hélice-coude-hélice (helix-turn-helix en anglais) relativement flexibles, disposés presque linéairement avec une rotation de
2 à 3° dans le sens inverse des aiguilles d'une montre.
Cet empilement de répétition ANK pourrait se se comporter comme un ressort réversible lorsqu'elle est étirée, et pourrait fonctionner dans un mécanisme de couplage dans des tissus comme le cœur (Nanospring behaviour of ankyrin repeats 2006)
Reconnaissance par
les protéines ANK
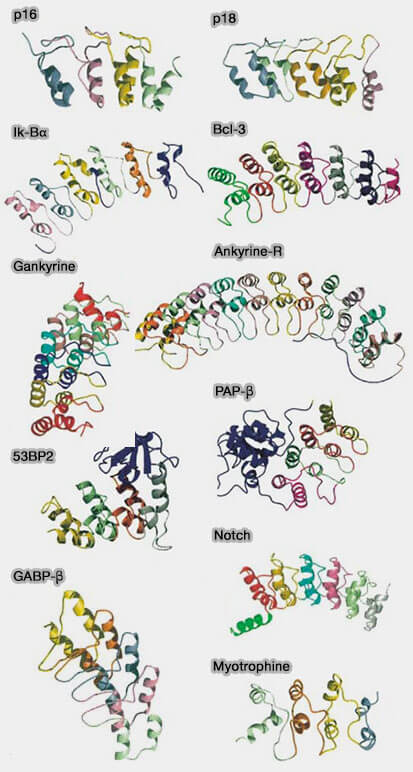
1. La structure des protéines contenant des répétitions ANK, classe de protéines de liaison la plus fréquente après les immunoglobulines, :
est stabilisée par les interactions de ces motifs,
a. La présence d'autres domaines dans la protéine affecte l'orientation des domaines ANK.
b. Le nombre de répétitions dans un domaine ANK, qui fournit une plateforme pour les interactions protéiques, est en de 4 à 7, mais ce nombre peut aller jusqu'à 29.
La superhélice de la protéine tourne d'environ 13° à chaque répétition (2-3° entre les répétitions.
Il en faudrait 32 pour faire un tour complet.
c. La variation de certains résidus de la répétition permet la spécificité des interactions moléculaires individuelles.
D'une part, les motifs ANK N- et C-terminaux d'une protéine, différents des motifs ANK internes, sont plus accessibles au solvant et protègent le noyau hydrophobe de son action.
D'autre part, le nombre de résidus de certains motifs peut diminuer (INK4, Inhibitor of cyclin-dependent kinase 4) et on peut même trouver des moitiés de motifs (GABP-α).
d. Les boucles reliant les différentes répétitions peuvent être plus ou moins longues.




 répétitions dans les protéines).
répétitions dans les protéines).