La sortie du cycle de l’APC/C implique la dissociation des E2, la dégradation des coactivateurs et la fermeture du module catalytique, assurant la transition ordonnée vers la phase G1.
Cette libération du substrat marque la fin d’un cycle catalytique et s’accompagne d’une réorganisation progressive du complexe APC/C vers son état inactif.
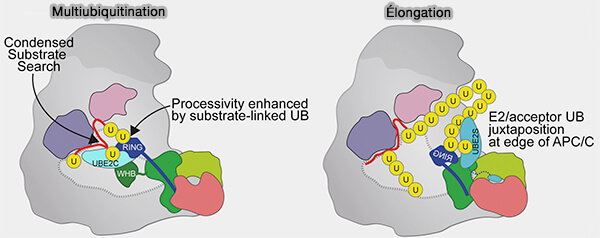
Multiubiquitination et élongation
(Figure : vetopsy.fr d'après Brown et coll)
Élongation de la chaîne et instabilité de l'APC/C
Lorsque la chaîne polyubiquitine atteint une longueur critique, la liaison du substrat devient instable, entraînant sa dissociation du module de co-reconnaissance APC10-coactivateur.
1. Au-delà de 6-8 unités d’ubiquitine, la chaîne devient volumineuse et hautement mobile, ce qui perturbe l’ancrage stable du substrat dans la poche de reconnaissance formée par le coactivateur (Cdc20/Cdh1) et APC10.
Une fois la chaîne polyubiquitine suffisamment longue, ces dégrons sont masqués par la topologie de la chaîne ou tirés hors du site par les mouvements de l’ubiquitine.
Le substrat glisse alors hors du site de reconnaissance.
Ce repliement réduit l’accessibilité du site actif et empêche toute nouvelle réaction avant la réactivation du complexe.
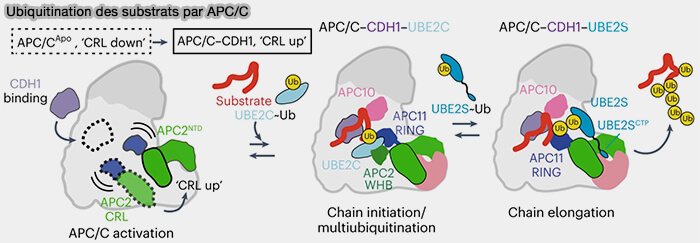
Ubiquitination des substrats par le complexe APC/C
(Figure : vetopsy.fr d'après Bodrug et coll)
Libération de Ube2S
Ube2S est libéré du domaine RING d’APC11, puis rechargé en ubiquitine pour un nouveau cycle.
1. Une fois la chaîne allongée et le substrat libéré, Ube2S se détache à son tour, la conformation du RING repassant à un état inactif.
a. En effet, le substrat, désormais porteur d’une chaîne polyubiquitine complète, perd son affinité pour le module de reconnaissance APC10-Cdh1/Cdc20 et se détache du complexe.
Dès lors, il n’existe plus de lysine accepteur correctement positionnée à proximité du site catalytique d’Ube2S.
Le site actif d’Ube2S (Cys95) ne peut plus transférer son ubiquitine, ce qui supprime la contrainte catalytique qui stabilisait l’enzyme dans sa position active.
b. Après la libération du substrat, le module APC2-APC11 subit une relaxation structurale.
Les surfaces d’interaction qui maintenaient Ube2S en place, via APC11 et l’ancrage CTP sur APC2-APC4/5, se désalignent.
c. La libération d’Ube2S a aussi un rôle préventif.
Si elle restait fixée au RING d’APC11, son thioester E2~Ub pourrait réagir avec d’autres lysines du complexe lui-même et provoquer une autoubiquitination non spécifique.
Le repliement du module et la dissociation d’Ube2S protègent le complexe de ces réactions parasites.
Ce renouvellement rapide des E2 permet à l’APC/C de fonctionner en rafales catalytiques, i.e. burst cycles, alternant entre phases actives et périodes d’inactivité conformationnelle.
Libération des coactivateurs
Le coactivateur (Cdc20 ou Cdh1) demeure transitoirement associé à l’APC/C à la fin du cycle catalytique, avant d’être remplacé ou inactivé selon la phase du cycle cellulaire.
Cette alternance entre Cdc20 et Cdh1 constitue un mécanisme de reprogrammation temporelle garantissant la directionnalité de la progression mitotique.
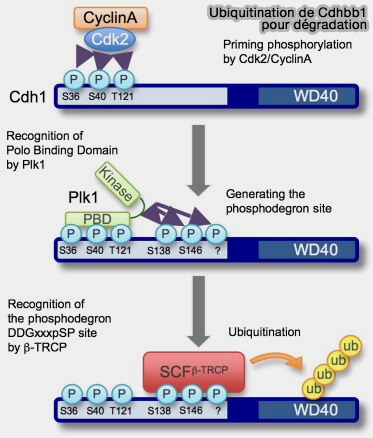
Ubiquitination de Cdh1 pour sa dégradation
(Figure : vetopsy.fr d'après Fukushima et coll)
L’APC/CCdc20 catalyse sa polyubiquitination autorégulée, entraînant sa dégradation par le protéasome 26S et provoquant la transition vers l’état APC/CCdh1.
Ce basculement coïncide avec la chute de l’activité CDK1, qui permet la déphosphorylation de Cdh1 et son recrutement sur la plateforme APC/C.
2. Durant la phase G1, APC/CCdh1 maintient le repos du cycle en éliminant les restes de substrats mitotiques, i.e. sécurine, cycline B, géminine…
Cette double régulation, i.e. inhibition par EMI1 et dégradation par SCFβ-TrCP, assure la désactivation complète de l’APC/C avant la réplication de l’ADN.
2. Le retour à l’état basal s’accompagne d’une restructuration conformationnelle du cœur de l’APC/C, notamment au niveau des TPR et du module APC2-APC11.
Cette réinitialisation maintient le complexe dans une forme " fermée " inerte, prête à être réactivée lors du cycle suivant.
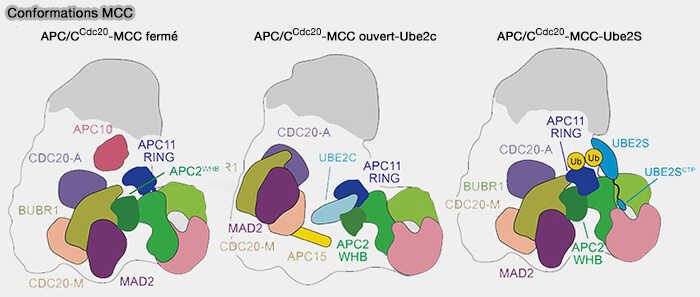
Conformations du MCC
(Figure : vetopsy.fr d'après Bodrug et coll)
L’ensemble du processus illustre la capacité de l’APC/C à s’autoréguler par couplage entre catalyse et dégradation de ses propres cofacteurs, garantissant la directionnalité et l’irréversibilité de la progression du cycle cellulaire, et assurant la transition ordonnée entre mitose, sortie de mitose et phase G1.




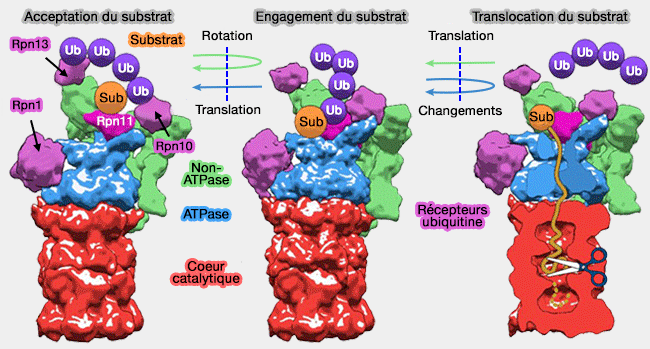
 mécanisme du protéasome).
mécanisme du protéasome).