Des modes spécialisés de nucléation non centrosomale à l'appareil de Golgi, au cortex et dans les neurones adaptent l’organisation des microtubules aux exigences de la polarité et de la différenciation cellulaire.
Nucléation au niveau du Golgi
L'appareil de Golgi constitue un site majeur de nucléation non centrosomale dans les cellules polarisées, en particulier lorsque l’organisation du réseau microtubulaire doit être orientée et asymétrique (The centrosome-Golgi apparatus nexus 2014).
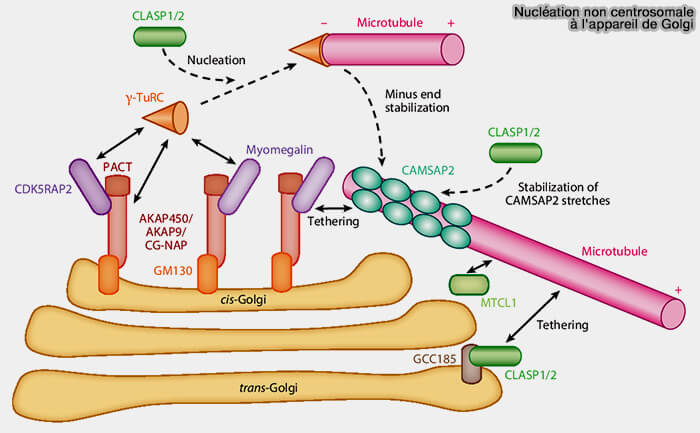
1. L’activité nucléatrice du Golgi repose sur le recrutement localisé du γ-TuRC sur un module d’ancrage spécifique associé aux membranes golgiennes, sans organisation radiale, distincte du modèle centrosomal (Microtubule-Organizing Centers 2017).
GM130 joue un rôle d’adaptateur structurant, organisant et stabilisant les sites d’ancrage du complexe nucléateur au sein des citernes golgiennes.
AKAP450 et les protéines apparentées au Golgi (CDK5RAP2-like/Myomegalin) partagent un domaine PACT impliqué dans l’ancrage du γ-TuRC aux plateformes de nucléation à la surface du Golgi et définissant des points de nucléation spatialement contraints.
CLASP, en coopération avec GM130, stabilise les protofilaments naissants, limite les événements de catastrophe et favorise l’établissement de microtubules persistants à partir du Golgi.
Nucléation non centrosomale au niveau de l'appareil de Golgi
(Figure : vetopsy.fr d'après Wu et Akhmanova)
Remarque : certaines protéines associées au Golgi apparaissent sur la figure car elles conditionnent la géométrie, la stabilité et la polarité du réseau microtubulaire golgien, sans constituer des facteurs de nucléation au sens strict.
Les golgines du trans-Golgi, tels que GCC185, participent à l’ancrage des extrémités (-) et à l’orientation des microtubules nouvellement formés vers la périphérie cellulaire.
CLASP, en coopération avec GM130 et GCC185, stabilise les protofilaments naissants, limite les événements de catastrophe et favorise l’établissement de microtubules persistants à partir du Golgi.
D’autres facteurs comme CAMSAP2/patronine stabilisent les extrémités (-) des microtubules après leur émergence, tandis que des protéines de stabilisation telles que CLASP limitent la catastrophe et favorisent la persistance des microtubules naissants.
Le complexe MTC1, associé au Golgi, intervient dans l’organisation et la stabilisation du réseau microtubulaire golgien, en contribuant à l’arrimage des microtubules et à la cohérence de leur orientation.
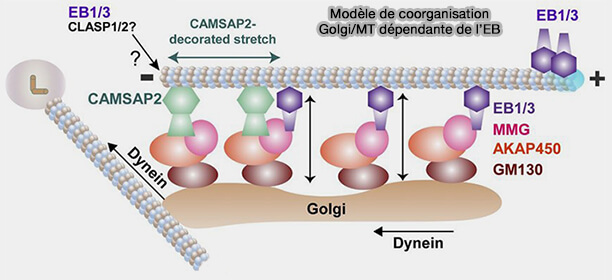
2. Sur certaines figures de synthèse, des +TIPs comme EB1 et EB3 sont représentés au Golgi via leur interaction avec des adaptateurs comme Myomegalin (MMG) qui s’associent à AKAP450 et GM130.
Ils contribuent à la régulation de la dynamique et de l’organisation des microtubules, en particulier en modulant l’extension des segments décorés par CAMSAP2 et la coopérative interaction des extrémités (-) avec le réseau golgien (Molecular Pathway of Microtubule Organization at the Golgi Apparatus 2016).
Modèle de coorganisation Golgi/MT dépendante de l’EB
(Figure : vetopsy.fr d'après Wu et coll)
3. La nucléation golgienne contribue à plusieurs fonctions cellulaires clés.
Elle génère un réseau microtubulaire anisotrope, orienté préférentiellement vers la périphérie cellulaire.
Elle soutient le transport vésiculaire directionnel et l’architecture de la voie sécrétoire.
Elle participe au maintien et au renforcement de la polarité cellulaire, notamment dans les cellules épithéliales et migratrices.
Elle peut suppléer partiellement l’activité centrosomale, voire devenir dominante selon le type cellulaire et l’état de différenciation.
Nucléation dépendante du cortex cellulaire
Le cortex cellulaire peut servir de plateforme de nucléation localisée, permettant de générer des microtubules indépendants du centrosome selon deux mécanismes principaux.
CAMSAP/patronine et stabilisation des extrémités (-)
La nucléation dépendante du cortex repose principalement sur la stabilisation précoce des extrémité (-) des microtubules par les protéines de la famille CAMSAP/patronine/Nezha, et non sur la création d’un noyau canonique via les complexes γ-TuRC.
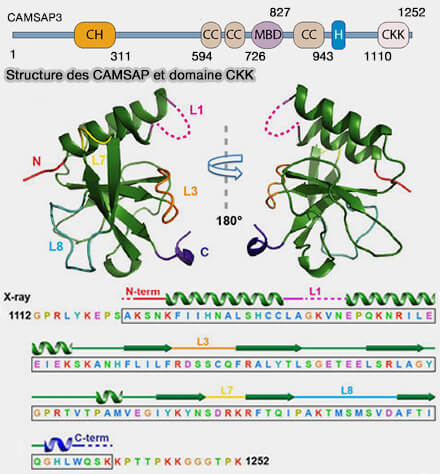
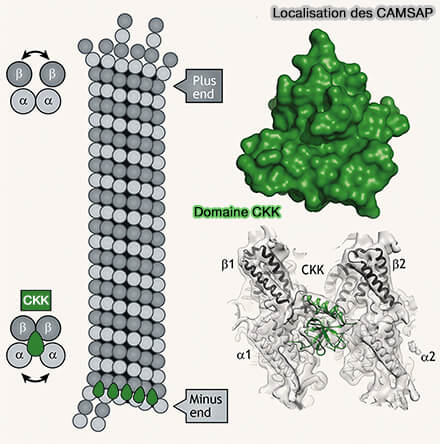
Structure de CAMSAP3 et domaine CKK
(Figure : vetopsy.fr d'après Aterthon et coll et autres sources)
Remarque : CAMSAP (Calmodulin-regulated Spectrin-Associated Protein) désigne une famille de protéines conservées, dont la patronine est l’homologue de la drosophile, et Nezha le nom historique de CAMSAP3.
Chez l’homme, on trouve trois protéines paralogues, CAMSAP1, CAMSAP2, appelé également CAMSAP1L1/KIAA1078 et CAMSAP3, aussi connu sous le nom de Nezha/KIAA1543.
a. Le domaine CH (Calponin Homology) N-terminal est impliqué dans des interactions protéine-protéine et la localisation subcellulaire.
Un domaine MBD (Microtubule-Binding Domain), présent uniquement chez CAMSAP2 et CAMSAP3/Nezha, localisé entre CC2 et CC3, contribue à la liaison latérale au lattice du microtubule, mais ne reconnaît pas l’extrémité (-).
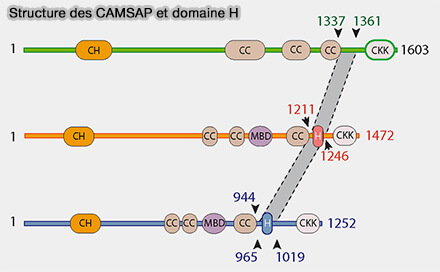
Structure des CAMSAP et domaine H
(Figure : vetopsy.fr d'après Li et coll) Un domaine hélicoïdal (H) situé entre CC3 et le domaine CKK, est conservé chez tous les CAMSAP.
Le domaine H (α-hélice) des CAMSAP ne médie pas la liaison au microtubule à lui seul, mais confère la spécificité de reconnaissance du minus end, en modulant le mode d’interaction du domaine CKK avec le lattice proximal.
Le comportement vis-à-vis des extrémités (-) diffère selon les membres de la famille.
1. CAMSAP1 s’associe de manière transitoire aux extrémités (-) et les suit, sans stabilisation durable du microtubule.
CAMSAP1 se fixe à l'extrémité (-) naissante, mais ne forme pas une coiffe protectrice stable.
Elle accompagne l’extrémité (-) lorsqu’elle est transitoirement exposée ou en croissance lente, puis se dissocie, contrairement à CAMSAP2 et CAMSAP3 qui verrouillent et stabilisent durablement cette extrémité.
Remarque : la croissance des microtubules est majoritairement portée par l’extrémité (+), la croissance de l’extrémité (+) ne survient que dans des contextes non centrosomaux spécifiques.
Localisation des CAMSAP
(Figure : vetopsy.fr d'après Akhamova et coll)
a. Cette interaction forme une coiffe protéique stable qui empêche l’accès des facteurs de dépolymérisation, i.e. enzymes de coupure AAA+, et bloque la perte rapide de dimères α/β-tubuline.
b. La fixation de CAMSAP2/3 :
stabilise des protofilaments courts et intrinsèquement instables, en abaissant la barrière énergétique de l’initiation,
inhibe les catastrophes et bloque la dépolymérisation des extrémités (-),
transforme des noyaux transitoires en microtubules capables d’entrer en phase d’élongation.
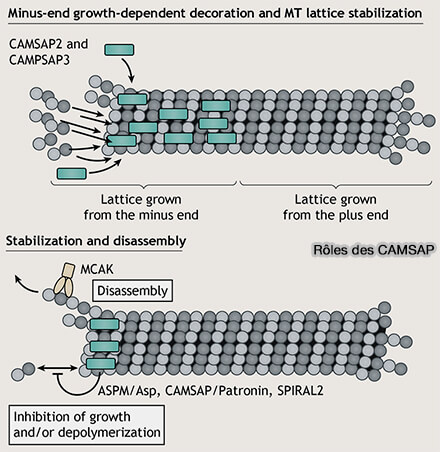
Les CAMSAP ne nucléent pas les microtubules, mais sécurisent l’existence des extrémités (-) des polymères naissants, transformant des structures instables en éléments durables du réseau cortical.
Contrairement à la nucléation centrosomale par les complexes γ-TuRC, ce mécanisme :
Rôles des CAMSAP
(Figure : vetopsy.fr d'après Akhamova et coll)
ne repose pas sur un gabarit géométrique,
ne définit pas une polarité radiale globale,
produit des microtubules ancrés localement par leur extrémité (-), laissant l’extrémité (+) libre pour une croissance orientée.
Le réseau ainsi formé est non radial, spatialement contraint par le cortex, et s’organise en faisceaux ou segments orientés en fonction des besoins locaux de la cellule.
Remarque : au cortex cellulaire, les CLASP s’associent au lattice proximal des microtubules en croissance et sont recrutées sur des plateformes membranaires telles que LL5α/β.
Ce module ne participe pas à la nucléation, mais limite la catastrophe, favorise des pauses ou des rescues locaux et permet l’établissement de microtubules corticaux persistants, orientés selon les contraintes de la membrane.
ASPM (Spiral chez Drosophila) est une MAP du pôle du fuseau mitotique impliquée dans la focalisation des extrémités (−) des microtubules mitotiques.
Rôles cellulaires des CAMSAP
Cellules épithéliales et cellules migratrices
Dans les cellules épithéliales et migratrices, cette nucléation corticale permet :
l’établissement de réseaux cortico-centraux alignés avec l’axe de polarité cellulaire,
l’orientation préférentielle des microtubules vers des domaines membranaires spécifiques,
l'adaptation fine de l’architecture microtubulaire aux contraintes locales de forme, d’adhérence et de mouvement.
Ce mode de nucléation soutient une organisation spatiale du réseau compatible avec la polarité, la migration dirigée et la compartimentation fonctionnelle de la membrane.
Neurones et cellules très polarisées
Dans les neurones et, plus généralement, dans les cellules à géométrie fortement polarisée, la nucléation non centrosomale devient le mode principal de production de nouveaux microtubules, le centrosome perdant progressivement son rôle de MTOC dominant.
1. La nucléation se déroule directement au sein du réseau existant selon plusieurs mécanismes complémentaires :
stabilisation de courts segments par CAMSAP/patronine, assurant une croissance orientée à partir des extrémités (-),
protection des microtubules contre les contraintes thermiques et mécaniques par MAP6,
amplification de la croissance par XMAP215/ch-TOG, à partir des microtubules préexistants, par élongation accélérée des polymères naissants, issus soit d’une nucléation par ramification, soit de la stabilisation de courts segments,
fragmentation contrôlée par les enzymes de coupure AAA+, i.e. katanine, spastine, fidgetine, générant des segments courts conservant leur polarité (+)/(-) et pouvant s’insérer dans un réseau existant ou amorcer un nouveau faisceau.
2. Dans les axones et dendrites, cette nucléation non centrosomale :
soutient l’allongement progressif de l’axone, en fournissant des microtubulses additionnels loin du corps cellulaire,
maintient la polarité interne des réseaux dendritiques et axonaux,
permet la réparation locale après contraintes mécaniques,
fournit une plasticité structurale indispensable dans les neurones matures, où le centrosome perd sa fonction de MTOC dominant.
Rôle transversal de XMAP215/ch-TOG
XMAP215/ch-TOG (Colonic and Hepatic Tumor Overexpressed Gene) n’est pas un nucléateur autonome, mais le principal facteur d’élongation associé aux microtubules naissants.