


Neurophysiologie : synapse
Vésicules synaptiques : cycle vésiculaire
Recyclage vésiculaire
: 6. Endocytose
Endocytose de masse dépendante de l'activité (ADBE)
Formation des vésicules synaptiques (VS) à partir du " bulk endosome "
- Neurophysiologie
- Cellules gliales et névroglie
- Neurones
- Potentiels membranaires
- Circuits neuronaux
- Neurotransmetteurs
- Récepteurs membranaires
- Synapses
{kind=link}

La formation des vésicules synaptiques à partir des " bulk endosomes " implique un grand nombre de molécules dont certaines interviennent déjà dans sa formation.
Vue d'ensemble de la régénération des VS
Une fois les " bulk endosomes " formés, ils doivent générer des vésicules synaptiques (VS), processus qui n'est pas dépendant de l'activité, pour reconstituer le pool de réserve des VS.
On note une absence de génération accélérée de VS à partir d'endosomes ou de disponibilité de VS dérivées d'endosomes massifs pour le réapprovisionnement du pool de réserve (RP) pendant les trains répétés de potentiels d'action.
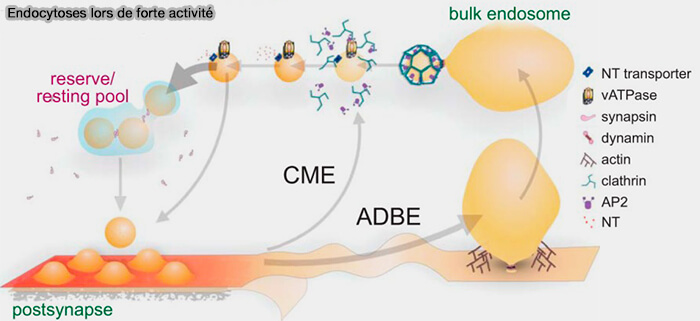
(Figure : vetopsy.fr d'après Chanadey et coll)
1. La régénération des VS, sous le contrôle de la clathrine, se produit au niveau de l'endosome internalisé, plutôt qu'à la membrane.
Dans l'ADBE et l'UFE, les endosomes sont formés directement à partir de la membrane plasmique.
- La formation d'endosomes peut se produire à différentes échelles de temps, i.e. dizaines de millisecondes pour l'UFE, à des dizaines de secondes, en fonction de la force de l'apport de stimulus.
- L'assemblage relativement lent de la couche de clathrine signifie que le bourgeonnement est finalisé sur l'endosome plutôt qu'à la membrane, en raison de l'invagination rapide de ces structures.
2. La régénération des VS à partir nécessite (Synaptic vesicle generation from activity-dependent bulk endosomes requires calcium and calcineurin 2013) :
- l'efflux de calcium précédemment accumulé dans l'endosome,
- l'activation de la calcineurine.

Bien que la formation des VS ne soit pas dépendante de l'activité, elle est quand même dépendante du calcium et de la calcineurine (Synaptic Vesicle Generation from Central Nerve Terminal Endosomes 2015).
Premier temps : efflux du Ca++
Après l'ADBE, l'essor ultérieur de la membrane interne nécessite l'efflux du calcium extracellulaire précédemment accumulé, qui est entraîné par l'acidification endosomale (Synaptic Vesicle Generation from Activity-Dependent Bulk Endosomes Requires Calcium and Calcineurin 2013).
Vue d'ensemble
1. Dans un premier temps, le Ca++ extracellulaire s'accumule à l'intérieur des endosomes massifs dans la gamme millimolaire par absorption de phase fluide (Synaptic Vesicle Generation from Central Nerve Terminal Endosomes 2015).
- Les expériences utilisant soit des tampons de calcium intracellulaires appliqués après la génération d'endosomes massifs, i.e. pour chélater le calcium s'écoulant d'endosomes, soit un dextrane conjugué à des tampons de calcium, i.e. pour chélater le calcium à l'intérieur des endosomes, ont toutes deux entraîné un retard dans la génération des VS ((Synaptic vesicle generation from activity-dependent bulk endosomes requires calcium and calcineurin 2013).
- L'efflux de calcium dépend de l'acidification du " bulk endosome ", car, l'inhibition de cet événement après la production de l'endosome arrête la production des VS et la reconstitution du pool de VS.
2. L'efflux de calcium dépend de l'acidification du " bulk endosome ", i.e. processus similaire à celui de l'acidification des vésicules synaptiques.
a. Le Ca++ extracellulaire doit être évacué en même temps que l'endosome s'acidifie.
b. L'inhibition de la calcineurine, protéine phosphatase dépendante du calcium, entraîne l'arrêt du bourgeonnement des VS et la reconstitution du pool de réserve (RP) de VS, ce qui suggère qu'elle est activée par l'efflux de Ca++ provenant des " bulk endosomes ".
La délocalisation de la calcineurine des " bulk endosomes " à l'aide de peptides compétitifs imitant un motif d'accueil PxIxI perturbe le bourgeonnement du VS.
Mécanismes possibles
Le mécanisme moléculaire sous-jacent à l’efflux dépendant du pH du calcium du " bulk endosome " est encore obscur.
Le mécanisme le plus probable est la présence d'un canal qui dépend des ions Ca++ et détecte l’acidification massive de l’endosome.
1. Le canal TRPV2 pourrait être activé par un pH faible et présente des similitudes biophysiques avec un canal calcique de l'endosome dans les endosomes artificiellement agrandis de cellules HEK293 (Luminal Chloride-dependent Activation of Endosome Calcium Channels: patch clamp study of enlarged endosomes 2007).
2. Un autre candidat pourrait être la mucolipine-3/TRPML3.
- C'est un canal calcique exprimé de manière omniprésente et localisé dans Hrb-endosomes positifs et inactivés à des valeurs de pH inférieures à six (Mucolipin-3 Regulates Luminal Calcium, Acidification, and Membrane Fusion in the Endosomal Pathway 2011).
- L'inactivation de ce canal retarde considérablement l'efflux de calcium dépendant du pH des endosomes dans un système non neuronal, i.e. il pourrait remplir une fonction similaire dans les endosomes terminaux nerveux.
Sélection du cargo
Protéines adaptatrices
La sélection du cargo se produit très probablement pendant l'étape de l'acidification, étant donné que l'adaptateur de membrane plasmique classique AP-2 et l'AP-1/AP-3 endosomal sont nécessaires pour la génération des VS à partir d'endosomes massifs (Clathrin/AP-2 Mediate Synaptic Vesicle Reformation from Endosome-like Vacuoles but Are Not Essential for Membrane Retrieval at Central Synapses 2014 et Synaptic Vesicle Generation from Central Nerve Terminal Endosomes 2015).
Les adaptines (AP ou Adaptor Protein) sont une famille de complexes hétérotétramères apparentés qui fonctionnent comme des revêtements pour le tri des cargos de protéines membranaires intégrales dans le système endomembranaire.
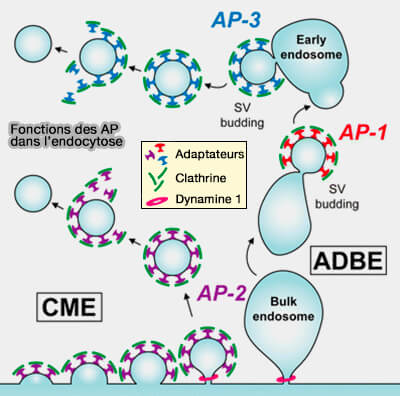
(Figure : vetopsy.fr d'après Cheung et coll)
1. L'AP-1 et l'AP-3 sont des protéines adaptatrices dans l'ADBE (Adaptor Protein Complexes 1 and 3 Are Essential for Generation of Synaptic Vesicles from Activity-Dependent Bulk Endosomes 2012).
a. AP-1 et AP-3, localiss aux terminaux centraux, se sont révélées nécessaires uniquement pour le recyclage des VS au cours d'une activité intense.
- La brefeldine A, un métabolite fongique qui bloque le recrutement de l'AP-1 et de l'AP-3 dans les membranes en interférant avec la petite GTPase Arf1, a arrêté la génération de VS à partir des " bulk endosomes ", de même que l'inactivation du shRNA des sous-unités d'AP-1 et AP-3.
- Les souris AP-3b2 knockout présentent des déficits significatifs en libération asynchrone, soutenant le rôle essentiel de la protéine adaptatrice AP-3 dans la génération de VS à partir d'endosomes massifs (Vesicles derived via AP-3 dependent recycling contribute to asynchronous release and influence information transfer 2014).
b. La double exigence en AP-1 et AP-3 pourrait s'expliquer par la circulation du cargo via un endosome précoce après avoir quitté le " bulk endosome " (AP-1/σ1B-Dependent VS Protein Recycling Is Regulated in Early Endosomes and Is Coupled to AP-2 Endocytosis 2015).
En outre, le trafic des cargos des VS pourrait impliquer des intermédiaires endosomaux qui sont distincts des " bulk endosomes " (Loss of Skywalker Reveals Synaptic Endosomes as Sorting Stations for Synaptic Vesicle Proteins 2011).

Le tri des cargos pourrait s'effectuer via une relation complexe entre les compartiments endosomaux formés directement à partir de la surface cellulaire, i.e. par l'ADBE et l'UFE, et le système de recyclage qui partageraient certains processus.
2. Par contre, l'AP-2 ne serait pas indispensable dans l'ADBE, alors qu'elle l'est dans l'endocytose clathrine-dépendante (AP-1/σ1B-Dependent VS Protein Recycling Is Regulated in Early Endosomes and Is Coupled to AP-2 Endocytosis 2015 et Clathrin/AP-2 Mediate Synaptic Vesicle Reformation from Endosome-like Vacuoles but Are Not Essential for Membrane Retrieval at Central Synapses 2014).
Dans l'endocytose clathrine-dépendante (CME), les cargos des VS sont triés et groupés par l'AP-2 pour être récupérés au niveau de la membrane plasmique, ce qui permet la formation directe de VS à partir de ce compartiment.
3. Les molécules adaptatrices AP180/CALM, qui sont, avec AP-2, les deux autres protéines adaptatrices majeures de la CME, peuvent se lier au motif SNARE de VAMP4 (The Biochemical Properties and Functions of CALM and AP180 in Clathrin Mediated Endocytosis 2014).
CALM et AP180 garantissent que les VAMP soient triées en vésicules naissantes par liaison directe au domaine ANTH.
4. L'exigence de différentes protéines adaptatrices peut conduire à des VS avec des compositions moléculaires variables, ce qui entraîne l'hétérogénéité fonctionnelle (Synaptic Vesicle Recycling Pathway Determines Neurotransmitter Content and Release Properties 2019).
Cependant Il est tout à fait possible qu'aucun tri n'intervienne pendant l'ADBE, car tous les cargos des VS présents sur la membrane plasmique peuvent s'accumuler, même si l'absorption est non sélective.
Par conséquent, des questions fondamentales subsistent quant à la base moléculaire de la sélection des cargos des VS pour les " bulk endosomes ".
Adaptateurs et recrutement de la clathrine
En plus du regroupement de cargo, la plupart des adaptateurs recrutent également les protéines qui conduisent à l'assemblage de la protéine de clathrine.
- La génération d'endosomes par ADBE ou par endocytose ultrarapide (UFE) est indépendante de la clathrine chez les mammifères et les invertébrés, alors qu'elles sont dépendantes de la clathrine dans les terminaisons nerveuses des invertébrés.
- Cependant, la clathrine est indispensable pour la production de vésicules médiée par l'AP-3 à partir d'endosomes dans les lignées de cellules neuroendocrines (Clathrin regenerates synaptic vesicles from endosomes 2015).
Recrutement de PI(4,5)P2
1. Le recrutement de PI(4,5)P2 sur des domaines membranaires recrute les protéines adaptatrices car les endosomes massifs peuvent partager une composition très similaire à la membrane plasmique.
Le PI(4,5)P2 est généralement rapidement dégradé sur des membranes endocytaires hautement incurvées. Cependant, les structures de plus grand diamètre contiennent une quantité beaucoup plus élevée de ce phospholipide (Synaptojanin 1-Mediated PI(4,5)P2 Hydrolysis Is Modulated by Membrane Curvature and Facilitates Membrane Fission 2011).
Ainsi, la courbure peu profonde des " bulk endosomes ", combinée au contrôle calcium/calcineurine-dépendante de son métabolisme, peut être essentielle pour générer un pool labile de PI(4,5)P2 endosomal pour déclencher le recrutement des protéines d'adaptateur.
2. D'autres substrats de la calcineurine sont tous nécessaires pour une endocytose clathrine-dépendante (CME) efficace :
- la dynamine et la syndapine,
- les protéines de courbure comme l'AP180, l'epsine, l'Esp15 et l'amphiphysine.
Deuxième temps : endocytose CME
La régénération des VS, sous le contrôle de la clathrine, se produit au niveau de l'endosome internalisé, plutôt qu'à la membrane.
L'endocytose clathrine-dépendante (CME) est étudiée dans un chapitre spécial et les mécanismes de régénération des vésicules synaptique à partir du " bulk endosome " sont approximativement similaires.
Protéines adaptatrices
Rappelons que les protéines adaptatrices qui contribuent à la formation de la cage de clathrines sont l'AP-1 et l'AP-3, ce qui diffère de la CME classique qui utilise l'AP-2 ( cf. plus haut).
cf. plus haut).
Rôles de la dynamine ou/et des EHD
Quel que soit le statut de phosphorylation de la dynamine 1, son activité GTPase fait partie intégrante du bourgeonnement de la vésicule synaptique (VS) à part entière.
1. Le fait que la génération de VS n'ait pas été complètement supprimée dans le cas de neurones KO à la dynamine suggère que :
- soit des voies alternatives de génération de VS sont possibles en l'absence de dynamine,
- soit que la dynamine n'agit que pour faciliter le processus et n'est pas obligatoire pour l'événement.
Si la dynamine n’est pas suffisante ou nécessaire pour assurer la fission des VS à partir des " bulk endosomes ", la famille des protéines EHD (Eps15 Homology Domain ATPase), i.e. EHD1-4, pourrait jouer ce rôle.
2. Les protéines EHD (Eps15 Homology Domain ATPase), i.e. EHD1-4 protéines sont liées à la superfamille de la dynamine (DSP) et peuvent former des tubules à partir de liposomes, entraînant l'hydrolyse de l'ATP.
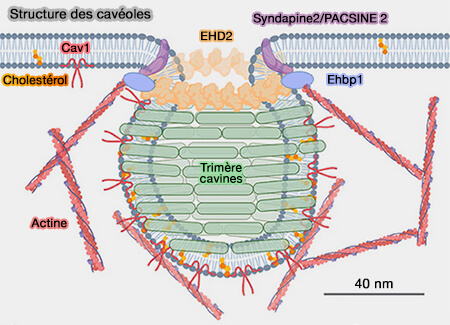
(Figure : vetopsy.fr d'après Matthaeus et coll)
a. L'EHD1 est concentrée in vivo au niveau des tubules membranaires qui sont enrichis en phospholipides PI(4)P et PI(4,5)P2, proposés comme étant présents sur les " bulk endosomes " (Eps15 Homology Domain 1-associated Tubules Contain Phosphatidylinositol-4-Phosphate and Phosphatidylinositol-(4,5)-Bisphosphate and Are Required for Efficient Recycling 2009).
b. L'EHD-1 interagit avec la calcineurine, substrat de l'amphiphysine-1 pour tubuler la membrane in vivo (AMPH-1/Amphiphysin/Bin1 functions with RME-1/Ehd in endocytic recycling 2010).
Dans ces études, la suppression de l'amphiphysine ou de l'EHD-1 a entraîné un défaut de trafic similaire au niveau du recyclage des endosomes chez C. elegans et également une mauvaise localisation de la syndapine.
c. Une interaction entre les membres de la famille EHD et la syndapine est nécessaire pour le bourgeonnement endosomal dans les cellules non neuronales.
Les EHD sont recrutés dans les endosomes via une interaction entre leurs domaines EH avec les motifs NPF de la syndapine (EHD Proteins Associate with Syndapin I and II and Such Interactions Play a Crucial Role in Endosomal Recycling 2005).
2. Ainsi, une déphosphorylation de la dynamine I dépendante de la calcineurine peut être nécessaire pour recruter la syndapine pour médier un événement de fission dépendant de l'EHD/amphiphysine qui est complètement indépendant de son activité GTPase.
Alternativement, les protéines EHD peuvent agir pour favoriser la génération de VS dépendante de la dynamine en contrôlant l'assemblage de l'hélice de dynamine (Regulation of Synaptic Vesicle Budding and Dynamin Function by an EHD ATPase 2011).
Déshabillage de la vésicule et regroupement dans le pool de réserve
1. Le processus de déshabillage de la cage est classique et dépend de :
- Hsc70/HSPA8 (70 kDa), protéine appartenant à la famille des protéines de choc thermique et des protéines chaperons ;
- l'auxiline/GAK, qui représente un membre des co-chaperons de Hsc70 (A sequential mechanism for clathrin cage disassembly by 70-kDa heat-shock cognate protein (Hsc70) and auxilin 2011).
Le déshabillage de la vésicule de clathrine est étudié dans un chapitre spécial.
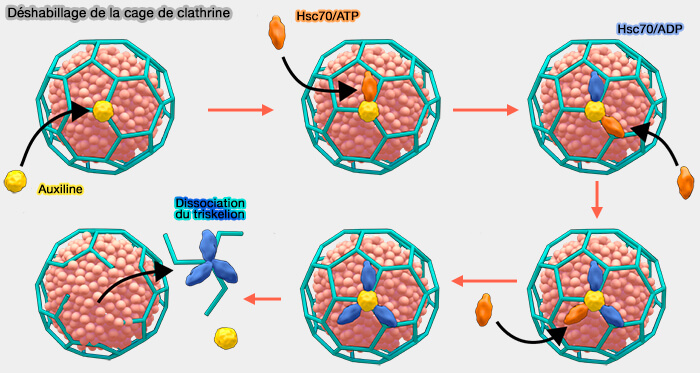
(Figure : vetopsy.fr d'après Mechanobiology institute de Singapour)
2. Après le déshabillage, les vésicules sont reproupées dans le pool de réserve, processus qui dépend de :
Le regroupement des vésicules est étudié dans un chapitre spécial.
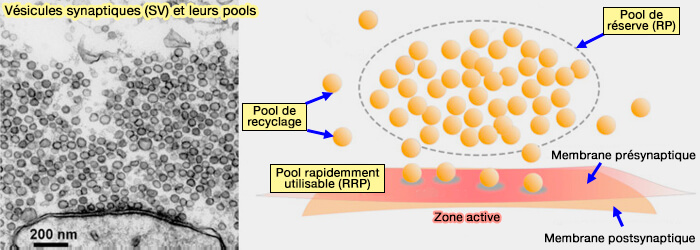
(Figure : vetopsy.fr d'après Zhang et coll)
Retour à l'ADBE
NeurophysiologeCellules gliales et névroglieNeuronesPotentiels membranairesCircuits neuronauxNeurotransmetteursRécepteurs membranairesSynapsesStructure synaptiqueNeurotransmissionVésicules synaptiques (VS)Pools synaptiquesRôles du calcium dans l'électrosécrétionCycle synaptiqueTranslocation et attache (tethering)Amarrage (docking) et amorçage (priming)Fusion et exocytoseRecyclage des VSDifférents types d'endocytoseAcidification et remplissage des VSRegroupement des VSPlasticité et régulation synaptique