lors de leur découverte, à la dénaturation thermique, d'où leur nom,
mais aussi au froid, aux UV ou pendant la cicatrisation des plaies ou le remodelage des tissus.
2. des protéines chaperons, i.e. protéines associées à la protection, au maintien et à la régulation des fonctions des protéines auxquelles elles sont associées.
Il existe d'autres protéines chaperons qui ne sont pas des protéines de choc thermique.
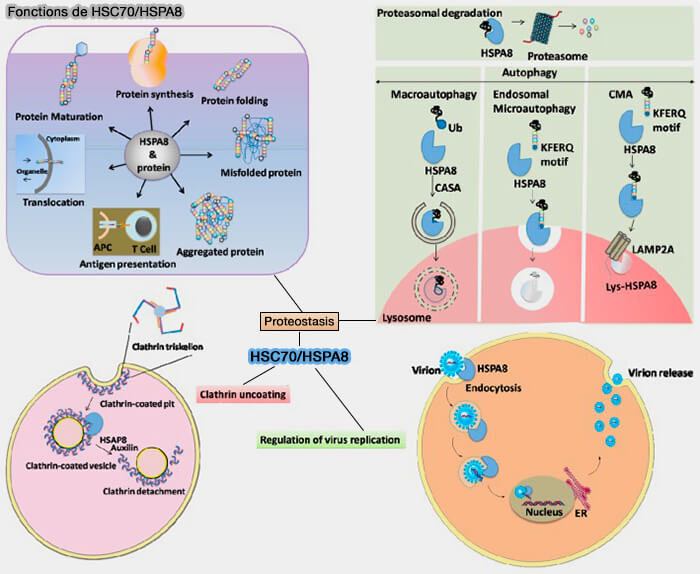
3. Hsc70/HSPA8 avec ses co-chaperons et ses chaperons constitue un réseau complexe et est associé à de nombreuses fonctions décisives pour la survie cellulaire, notamment le repliement des protéines et la dégradation des protéines, i.e.
régulateur de protéines, repliement, maturation, translocation, assemblage, démontage, agrégation, présentation et différenciation de l'antigène (réticulum endoplasmique et réponse UPR),
régulation de la réplication du virus (à la fois positif et négatif).
Fonctions de HSC70/HSPA8
(Figure : vetopsy.fr d'après Bonam et coll)
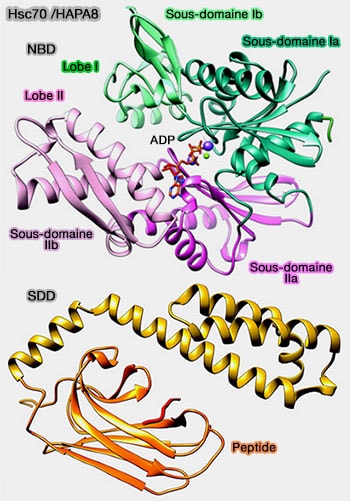
Structure de Hsc70
Hsc70 facilite le pliage des protéines pour les protéger, en cachant les acides aminés hydrophobes souvent impliqués dans les agrégations des protéines.
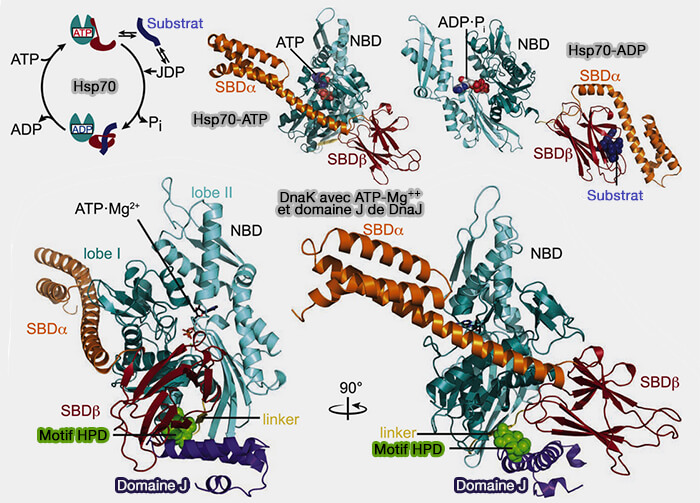
1. Le domaine N-terminal (NBD : Nucleotide Binding Domain 44kDa) est un domaine ATPase, constitué de 4 sous-domaines (IA, IB, IIA et IIB), divisé en deux lobes séparés par une profonde fente nucléotidique (comme dans toutes les ATPases).
L'hydrolyse de l'ATP en ADP et Pi conduit à des changements conformationnels des autres domaines.
2. Un lien (linker) est présent entre les deux domaines.
3. Le domaine de fixation au substrat (SBD : Substrate Binding Domain 24 kDa) est composé par :
SBD contient une fente qui peut accueillir des peptides de 5-7 acides aminés, en général hydrophobes, avec une préférence particulière pour la séquence FYQLALT.
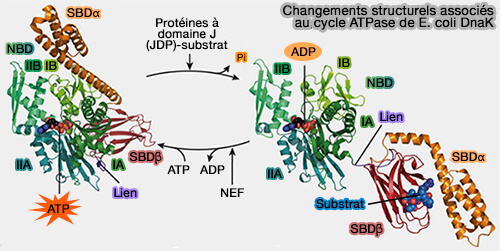
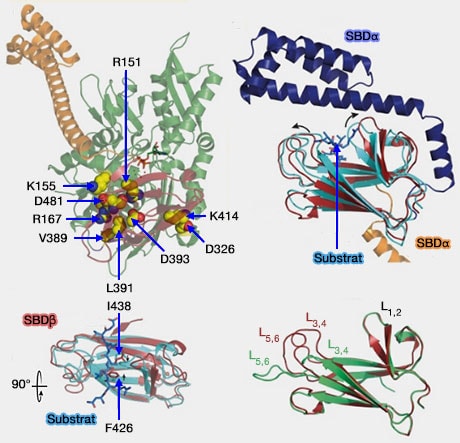
Changements structurels de DnaK de la levure
(Figure : vetopsy.fr d'après Kityk et coll)
b. un sous-domaine C-terminal, domaine constitué de plusieurs hélices α, qui stabilise la structure interne et agit comme un " couvercle " pour maintenir le substrat dans le tonneau.
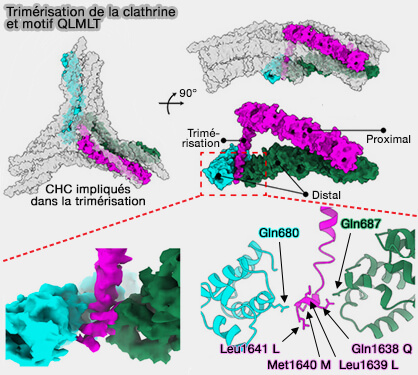
Trimérisation et motif QLMLT
(Figure : vetopsy.fr d'après Paran et coll)
a. Bien que les fonctions de liaison du NBD et du SBD soient séparables, l'activité de chaperon Hsp70 dépend strictement du couplage serré de ces deux domaines par la liaison de l'ATP, ce qui conduit à la modulation de leurs activités (Hsp70 chaperone dynamics and molecular mechanism 2013).
Hsc70/HSPA8
(Figure : vetopsy.fr d'après Mayer et Kityk)
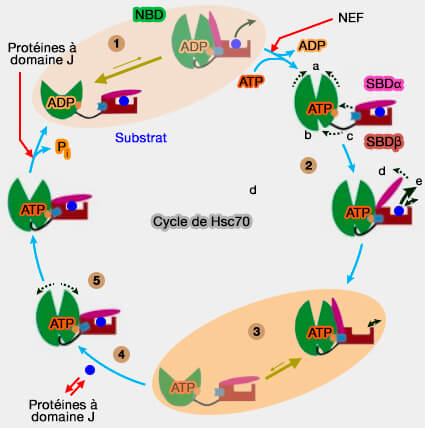
L'ATP réduit considérablement l'affinité des peptides pour les substrats peptidiques en accélérant à la fois les taux de libération et de liaison.
Dans le même temps, la liaison du peptide stimule la vitesse d'hydrolyse de l'ATP.
b. Grâce à ce couplage, l'énergie de l'hydrolyse de l'ATP est efficacement utilisée pour la régulation de la liaison et de la libération du peptide du substrat, i.e. le chaperon est efficace.
5. La liaison du substrat induit la fermeture du SBDα et la dissociation du SBDβ du NBD, ce qui permet la rotation des lobes NBD pour une hydrolyse optimale de l'ATP.
Les substrats stimulent l'hydrolyse de l'ATP à travers une voie distincte impliquant un déclenchement sur le NBD.




 réticulum endoplasmique et réponse UPR),
réticulum endoplasmique et réponse UPR),