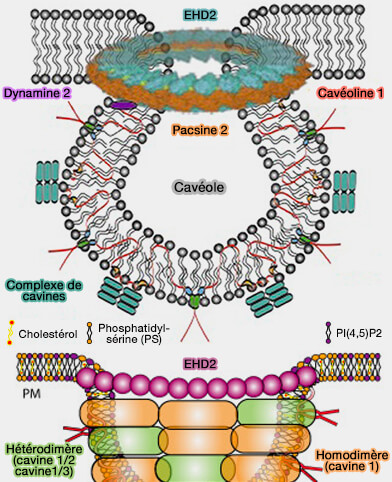
Les cavéoles, du latin caveolae, petites grottes, sont des invaginations de la membrane plasmique, en forme de Ω inversé, plus ou moins présentes selon le type cellulaire.
Les cavéoles sont de petites vésicules ovoïdes de 50 à 80 nm de diamètre, en forme de Ω inversé avec un cou (neck) appelé aussi entonnoir (funnel) ou pore.
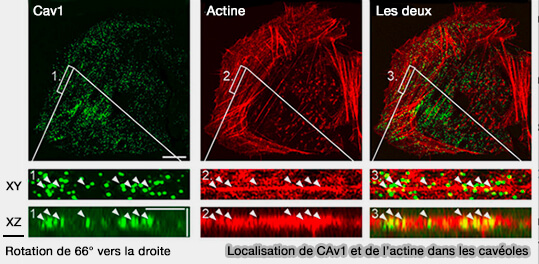
1. Cette morphologie permet facilement de les distinguer des cages de clathrine (100 nm), surtout que du côté intracellulaire, la membrane semble dépourvue de manteau protéique ou striée, contrairement aux cages de clathrine.
Leur cavité est reliée au milieu extracellulaire par le col, formant une sorte d'oméga (Ω) inversé.
Rosettes cavéolaires
(Figure : vetopsy.fr d'après Golani et coll)
La formation de ces invaginations a lieu au niveau des radeaux lipidiques (lipid raft). Les
cavéolines sont, de ce fait, utilisées
comme marqueurs moléculaires de ces structures.
une couche constituée par les extrémités N- et C-terminales des molécules de cavéoline (Cav) sous forme d'oligomères 8S organisée en une couche palmitoylée d'oligomères 70S, environ 160 Cav,
Composition des cavéoles
(Figure : vetopsy.fr d'après Lian et coll et Parton et coll)
une couche de cavines sous forme d'oligomères 60S, environ 50 cavines, disposée en spirale autour du manteau de cavéoline, i.e. les Cav et cavines forment un complexe 80S.
Remarque : des protéines transmembranaires comme ROR1 (Receptor tyrosine kinase-like Orphan Receptor 1), qui interagit avec CAV1, et la cavine1 pourraient faciliter la formation de cavéoles, mais pas dans tous les tissus.
L'assemblage des cavéoles dépend de leur composition lipidique spécifique et nécessite notamment la présence de cholestérol.
Les phospholipides chargés négativement les plus abondants de la membrane plasmique, i.e. la phosphatydylsérine (PS) et PI(4,5)P2 ou PIP2, se concentrent dans les cavéoles, formant des pools lipidiques physiologiquement distincts.
Cavéoles et phospholipides
(Figure : vetopsy.fr d'après Zhou et coll)
Les groupes de tête exposés au cytoplasme de ces phospholipides sont souvent impliqués dans l'association membranaire et la régulation allostérique des protéines associées à la membrane (Emerging Diversity in Lipid–Protein Interactions 2019).
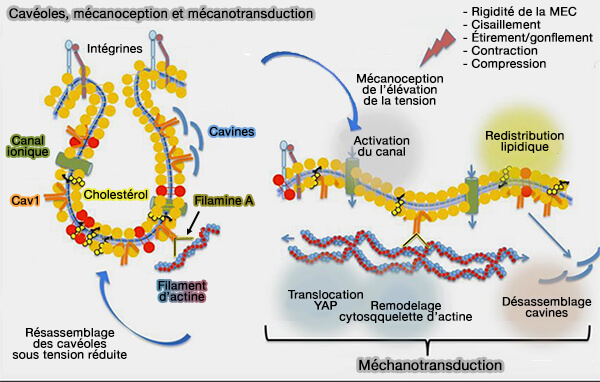
La déplétion en cholestérol déforme les cavéoles, ce qui implique que les PM ne conservent pas une morphologie invaginée sans cholestérol (The challenge of lipid rafts 2009).
La liaison de la syndapine 2/PACSINe2 à la membrane pourrait être régulée négativement par le cholestérol ( syndapine 2 et cholestérol).
Remarque : le domaine hélicoïdal de EHD2, i.e. α8-α12 (287-398), contient les sites de liaison membranaire.
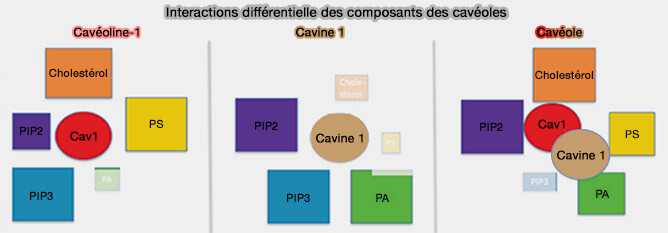
3. La composition lipidique, les protéines accessoires et l'architecture des domaines enrichis en cavéoline sont toutes impliquées dans l'association avec les cavines pour garantir que les cavéoles ne se forment qu'à la surface de la cellule ( modèle de biogenèse des cavéoles).
En effet, les liaisons lipidiques seules semblent cependant insuffisantes pour le recrutement des cavines à la surface membranaire, qui nécessite en outre la cavéoline-1.
Comme les cavéolines ne se lient pas aux cavines, excepté la cavine 3, plusieurs interactions de faible affinité pourraient être nécessaires pour générer le domaine cavéolaire.
L'assemblage ainsi que les interactions entre toutes ces molécules sont loin d'être connus.






 aplatissement des cavéoles) :
aplatissement des cavéoles) :