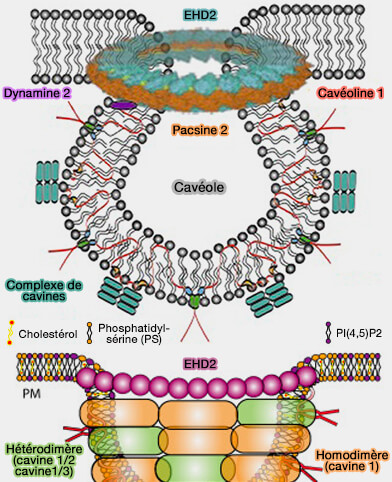
Les cavines sont nécessaires aux cavéolines oligomérisées pour former des cavéoles, en se trimérisant et en s'associant aux phospholipides membranaires, i.e. elles forment un complexe de revêtement externe qui remodèle la membrane en cavéoles..
1. La formation de la fosse cavéolaire implique l'action coordonnée de plusieurs protéines (Caveolae 2018).
a. Les protéines membranaires intégrales, i.e. les cavéolines (CAV), sont essentielles à leur formation, i.e. leur KO inhibent la genèse cavéolaire.
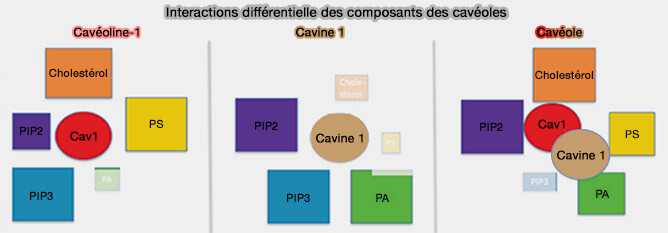
b. Les protéines de la membrane périphérique, i.e. les cavines, interagissent avec les cavéolines pour former un complexe de revêtement externe qui remodèle la membrane plasmique (MP) en cavéoles et les stabilisent par leurs interactions avec les lipides membranaires, i.e. cholestérol, phosphatidylsérine (PS), et PI(4,5)P2 ou PIP2.
c. Les complexes protéiques au niveau du cou impliquent :
Remarque : des protéines transmembranaires comme ROR1 (Receptor tyrosine kinase-like Orphan Receptor 1), qui interagit avec CAV1 et la cavine1, pourraient faciliter la formation de cavéoles, mais pas dans tous les tissus.
2. La composition lipidique de nanodomaines membranaires, en particulier, cholestérol et en phospholipides chargés négativement, i.e. phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2, est indispensable à la biogenèse des cavéoles.
L'assemblage ainsi que les interactions entre toutes ces molécules, i.e. protéines/lipides, protéines/protéines et lipides/lipides sont loin d'être connus, ce qui implique que certains articles un peu anciens ne sont plus d'actualité et que les modèles sont nombreux.
Vue d'ensemble des cavines
1. Les cavines sont nécessaires aux cavéolines oligomérisées pour former des cavéoles, en se trimérisant et en s'associant aux phospholipides membranaires, i.e. elles forment un complexe de revêtement externe qui remodèle la membrane en cavéoles.
Manteau des cavéoles (complexe 80S)
(Figure : vetopsy.fr d'après Ludwig et coll)
Les couches striées des cavéoles sont formées par les cavines polymérisées ( oligomérisation des cavines).
2. Quatre isoformes de la cavine ont été identifiées chez les mammifères, i.e. chaque membre de la famille a été initialement décrit dans des études les impliquant dans des processus cellulaires non apparentés.
Elle est suffisante, avec les cavéolines, pour générer des cavéoles en l'absence d'autres cavines.
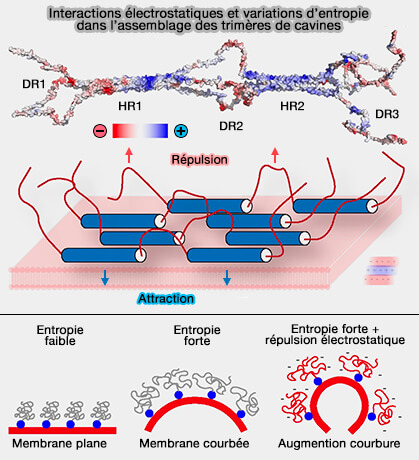
Électrostatisme et entropie dans l'assemblage des cavines
(Figure : vetopsy.fr d'après Tillu et coll)
Elle est nécessaire à l'association hétéromérique de la cavine 2 et la cavine 3.
b. La cavine 2, appelée aussi SDPR (Serum Deprivation-Response Protein) fut initialement découverte pour son abondance et sa haute affinité pour la phosphatidylsérine (PS) dans les plaquettes, puis décrite dans des expériences de déprivation. Elle est aussi exprimée dans les cellules endothéliales pulmonaires et le tissu adipeux.
c. La cavine 3, appelée aussi SRBC (Serum deprivation-response factor-related gene product that Binds to C-kinase), a d'abord été considérée comme l'adaptateur pour la protéine kinase Cδ (EC 2.7.10.2), d'où son autre nom PRKCDBP (Protein Kinase C Delta-Binding Protein).
Elle joue un rôle dans :
l'endocytose des cavéoles, i.e. la diminution de la cavine 3 diminue la dynamique de la cavéole,
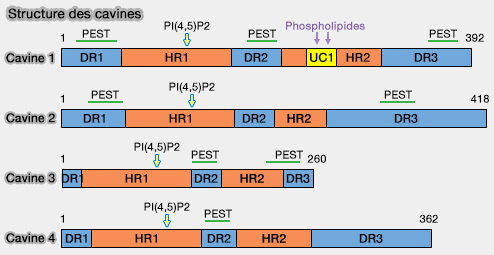
DR1 (Disordered Region 1) forme l'extrémité N-terminale.
DR1 contient des motifs PEST (séquences enrichies en proline, glutamate, sérine et thréonine) qui sont des sites de sensibilité protéolytique.
Cette région intervient dans l'association membranaire et le recrutement de CAV1( DR et oligomérisation).
HR1
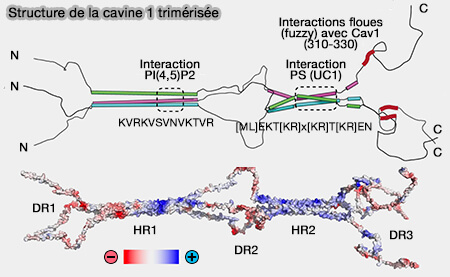
1. HR1 (Helical Region) est formée par une région α-hélicoïdale, dont la région centrale possède un domaine d'interaction, i.e. KVRKVSVNVKTVR, avec PI(4,5)P2 ou PIP2.
Cette région permettrait à la cavéoline de ne pas jouer le rôle de PTRF (Polymérase I and Transcript Release Factor), i.e. inhiberait ses séquences de localisation nucléaires (138-154 et 235-251)
DR2
DR2 (Disordered Region 2) suit HR1 et comporte des
motifs PEST.
HR2
1. HR2 (Helical Region) est formée par une région α-hélicoïdale, qui contiendraient plusieurs sites d'interaction avec la phosphatydylsérine (PS) sur HR2.
La séquence est la suivante : [ML]EKT[KR]x[KR]T[KR]EN.
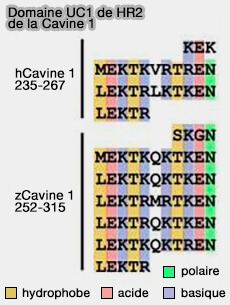
Les répétitions UC1 sont toujours insérées directement entre les séquences d'acides aminés hautement conservées KEK et LEKTR pointant vers la répétition de l'UC1
HR2 du poisson zèbre contient 5 UC1 et celui de des vertébrés terrestres seulement 2 seulement. Le nombre de répétitions UC1 est en corrélation avec la capacité des cavéoles à répondre à l'étirement de la membrane ( aplatissement des cavéoles).
La suppression du domaine UC1 dans la cavine 1 perturbe la formation de cavéoles.
La mutation des résidus potentiels de liaison membranaire dans le domaine UC1, i.e. les lysines (K) et les arginines (R), réduit l'affinité pour les membranes contenant de la phosphatidylsérine.
L'insertion de ces motifs dans la cavine 2, i.e. incapable de former des cavéoles, permet la formation des cavéoles.
DR3
DR3 (Disordered Region 2) suit HR2, forme la région C-terminale et contient, elle aussi, des motifs PEST.
Cette région intervient dans l'association membranaire et le recrutement de CAV1 ( DR et oligomérisation).
Remarques sur la structure des cavines
1. Les HR basiques sont bien conservées dans les cavines, i.e. HR1 et HR2 sont chargées positivement et ont donc une affinité pour les phospholipides chargés négativement.
HR1 montre une préférence pour les phosphoinositides comme PI(4,5)P2 ou PIP2.
2. Les domaines DR1, DR2 et DR3 ont une faible homologie de séquences, mais sont riches en chaînes latérales acides chargées négativement, ce qui à l'origine d'un modèle de remodelage membranaire ( oligomérisation des cavines).
Comme 15 et 20 trimères de cavines sont supposés être présents sur chaque cavéole et que chaque trimère possède au moins deux sites indépendants de liaison aux phospholipides, l'association membranaire et la polymérisation des cavines enrichissent localement la cavéole en PS et PI(4,5)P2 ou PIP2.





 oligomérisation des cavines).
oligomérisation des cavines).