Les cavines s'oligomérisent pour former des cavéoles avec les cavéolines et peuvent subir différentes modifications post-traductionnelles pour favoriser l'endocytose des cavéoles et leur dégradation.
Cavine 1/Cavine 2 et Cavine 1/Cavine 3 sont mutuellement exclus de ces complexes trimères de trimères et se retrouvent séparément à la surface des cavéoles.
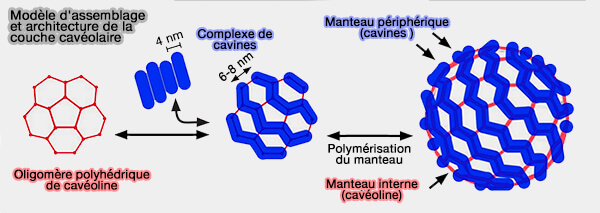
Une grande partie de chaque séquence cavine se replie en un assemblage de tiges très rigides et allongées (∼16 nm de longueur et ∼3 nm de largeur).
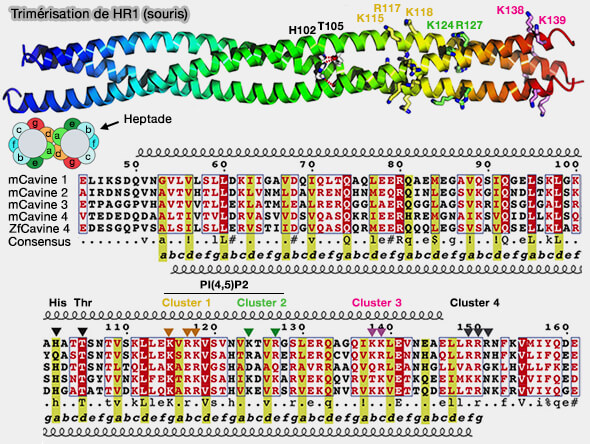
2. Le centre de HR1 possède une paire conservée de chaînes latérales histidine (H) et thréonine (T) liées à l'hydrogène, ce qui est rare dans les superhélices, et la cavine 2, elle, possède une glutamine (Q) à la place de l'histidine (H).
Cette thréonine peut être phosphorylée dans les cavines, bien qu'elle soit enfouie dans la structure.
D'une part, la phosphorylation de ce résidu perturbe l'appariement des liaisons hydrogène, et donc la stabilité de la structure, i.e. il est possible qu'elle représente un mécanisme de régulation de leur assemblage.
D'autre part, cette région contient également plusieurs sites d'ubiquitination, suggérant un niveau supplémentaire de régulation de l'assemblage et du renouvellement de la cavine.
Trimérisation de HR1 des cavines (souris)
(Figure : vetopsy.fr d'après Kovtun et coll)
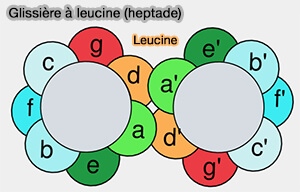
3. Les superhélices sont caractérisées par une répétition de motif hydrophobe de sept acides aminés, étiquetée abcdefg, appelée classiquement répétition heptade, qui définit les résidus centraux à l'interface entre les protomères qui favorisent leur oligomérisation ( heptade de la tropomyosine).
Les résidus désignés par les lettres a et d sont généralement hydrophobes et distribués en zigzag d'un côté de l'hélice en formant des bosses et des creux complémentaires qui s'emboîtent étroitement entre deux hélices dimérisées.
Glissière à leucine (heptade)
(Figure : vetopsy.fr)
Les séquences de ces heptades contribuent également à la spécificité différente des interactions cavine-cavine, mais on ne sait pas encore quels résidus sont cruciaux.
On trouve aussi plusieurs résidus glutamine (Q) conservés qui nécessitent un appariement à une chaîne latérale Q équivalente à partir de protomères adjacents, excluant ainsi les séquences de superhélices non apparentées d'interagir.
3. L'extrémité C-terminale de HR1 a une surface basique hautement conservée, composée d'amas de résidus lysine (K) et arginine (R), i.e. 145LLRRRN150 dans la cavine 1 qui sont sensibles à la phosphorylation.
Ces résidus pourraient jouer un rôle important dans l'association membranaire des cavines.
Si les DR ne sont pas indispensables pour la liaison membranaire, ils sont strictement nécessaires à la capacité de la cavine 1 de former des polymères capables de remodeler les membranes.
Les domaines désordonnés de cavine 1, en particulier le résidus 310-331 de DR3, s'associent avec la région désordonnée de CAV1, résidus 30 à 60 ( cf. plus bas).
Modèle d'assemblage et architecture de la couche cavéolaire
(Figure : vetopsy.fr d'après Ludwig et coll)
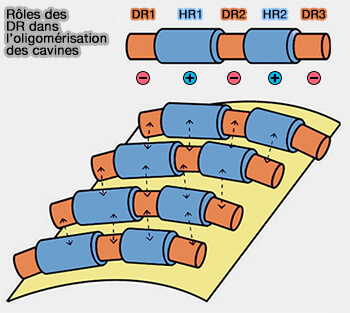
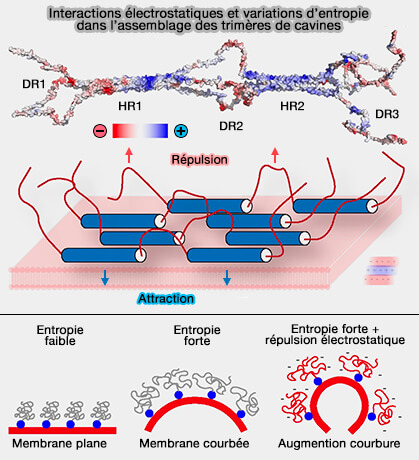
2. Les séquences HR sont basiques, i.e. chargées positivement alors que les DR (Disordered Region) sont acides, i.e. chargés négativement, ce qui a plusieurs conséquences.
a. D'une part, cette alternance de domaines permet aux DR d'un trimère de cavines de reconnaître les HR d'un trimère dans la strie parallèle adjacente et vice versa par des interactions électrostatiques transitoires par des LLPS ou Liquid–Liquid Phase Separation ( interactions floues et LLPS).
Rôles des DR dans l'oligoméristion des cavines
(Figure : vetopsy.fr d'après Kovtun et coll)
La cavine 1 purifiée peut facilement subir une LLPS dans des conditions proches de la physiologie et est capable de recruter CAV1 par le biais d'interactions impliquant la LLPS. Les domaines DR1 et DR3 contribuent à ce processus, bien qu'aucun domaine ne soit strictement essentiel.
Les mutations de DR3 qui maintiennent sa charge négative, mais empêchent l'interaction avec CAV1 au niveau de la membrane plasmique, favorisent la formation de gouttelettes liquides de cavine 1 dans les cellules.
L'élimination du domaine DR1 entraîne une formation de gel apparente plutôt qu'un assemblage de gouttelettes liquides in vitro, et dans les cellules, entraîne une accumulation de grandes structures intracellulaires qui contiennent également CAV1. Celles-ci sont formées par la redistribution endocytaire des structures de la cavéole à partir de la surface cellulaire et l'accumulation avec les membranes endosomales précoces.
b. D'autre part, les cavines seront en contact avec la membrane négative par les HR positifs et les DR négatifs seront repoussés vers l'extérieur.
Rôles des DR dans la courbure membranaire
Remarques préliminaires
1. Les deux manières de courber les membranes, en particulier dans l'endocytose clathrine-dépendante et dans d'autres vésiculations membranaires, i.e. qui nécessitent des motifs spécifiques, sont :
Ces motifs intrinsèquement désordonnés occupent des volumes considérablement plus importants par rapport aux motifs structurés de poids moléculaire équivalent et entraînent un encombrement stérique ( crowding et courbure/fission).
3. La liaison de telles protéines aux surfaces membranaires a deux conséquences majeures.
a. D'une part, la liaison d'une protéine désordonnée à une surface membranaire réduit considérablement l'entropie conformationnelle de la protéine.
Au fur et à mesure que la surface de la membrane s'éloigne de la protéine, i.e. prenant une forme convexe, l'entropie conformationnelle augmente.
Cette augmentation favorise la liaison des protéines aux surfaces membranaires courbes.
Ce processus est le principal moteur de détection de courbure dans le cas de domaines désordonnés longs de faible charge.
b. D'autre part, la charge négative de nombreux domaines désordonnés provoque une forte répulsion électrostatique par les lipides membranaires, qui ont généralement un fort potentiel de surface anionique (Membrane lipids: where they are and how they behave 2008).
L'augmentation de la courbure de la membrane augmente la séparation moyenne entre les acides aminés anioniques et les lipide et la répulsion électrostatique est minimisée lorsque les protéines désordonnées se lient à des surfaces fortement incurvées.
Ce mécanisme électrostatique est le principal moteur de la détection de courbure de domaines désordonnés courts de forte charge.
Ce mécanisme diffère fondamentalement des mécanismes précédents qui reposent sur des interactions électrostatiques attractives pour détecter la courbure de la membrane.
Applications aux cavines
Les séquences DR chargées négativement et les régions HR chargées positivement de la cavine 1 se combinent pour favoriser l'auto-association, l'interaction membranaire et l'encombrement des protéines à la surface de la membrane, entraînant une flexion ultérieure de la membrane.
Cependant, dans des conditions normales, le processus de génération de la courbure membranaire est étroitement régulé par CAV1, EHD2 et la syndapine 2/PACSINe 2, ainsi que par des lipides membranaires spécifiques, pour restreindre l'activité de remodelage de la cavine 1 uniquement aux cavéoles.
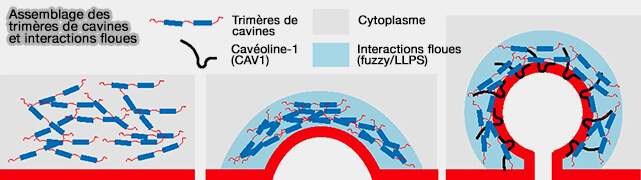
Assemblage des cavines et interactions floues (fuzzi)
(Figure : vetopsy.fr d'après Tillu et coll)
2. Ces interactions floues interviennent aussi dans le recrutement des cavines dans les domaines membranaires de CAV1, impliquant des interactions entre le domaine DR3 (310-331) et la région désordonnée de CAV1 (30 à 60).
Les altérations dans cette région de DR3 n'affectent pas la capacité de la cavine 1 de s'assembler en oligomères et de générer des tubules dans les membranes synthétiques, mais ne génèrent jamais de cavéoles.
Les cavines s'associent aux cavéolines dans des nanodomaines de la membrane plasmique pour former des cavéoles par des interactions floues d'affinité relativement faible, nécessitant une oligomérisation des cavines et des cavéolines ainsi que des interactions avec lipides membranaires ( biogenèse des cavéoles).
1. Les motifs PEST (séquences enrichies en proline, glutamate, sérine et thréonine) résidant dans les domaines DR des cavines sont un trait caractéristique de cette famille et des sites de sensibilité protéolytique.
L'abondance de ces sites suggère que la dégradation des protéines est un régulateur important de l'homéostasie et de la fonction de la cavine.
3. La phosphorylation implique des dizaines de sites identifiés expérimentalement répartis principalement dans les DR, sans signification fonctionnelle précise à ce jour.
Certains pensent que la phosphorylation de la cavine 1 pourrait être suffisante pour déclencher le désassemblage des cavéoles.





 biogenèse des cavéoles)
biogenèse des cavéoles)