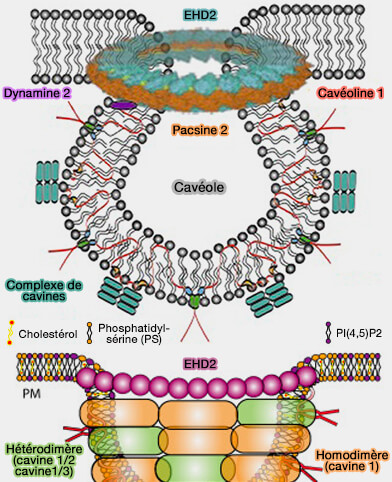
Les cavéoles, du latin caveolae, petites grottes, sont des invaginations de la membrane plasmique, en forme de Ω inversé, plus ou moins présentes selon le type cellulaire.
1. La formation de la fosse cavéolaire implique l'action coordonnée de plusieurs protéines (Caveolae 2018) :
a. Les protéines membranaires intégrales, i.e. les cavéolines (CAV), sont essentielles à leur formation, i.e. leur KO inhibe la genèse cavéolaire.
b. Les protéines de la membrane périphérique, i.e. les cavines, interagissent avec les cavéolines pour former un complexe de revêtement externe qui remodèle la membrane plasmique (MP) en cavéoles et les stabilisent par leurs interactions avec les lipides membranaires, i.e. cholestérol, phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2.
c. Les complexes protéiques au niveau du cou impliquent :
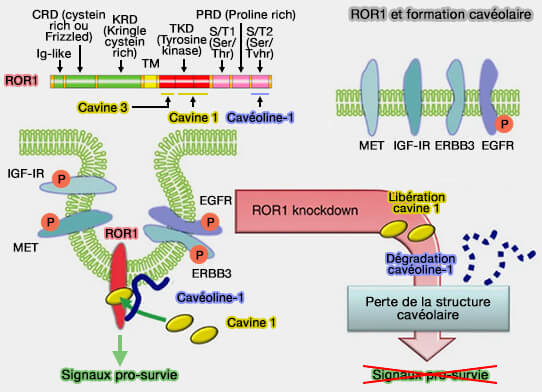
Remarque : des protéines transmembranaires comme ROR1 (Receptor tyrosine kinase-like Orphan Receptor 1), qui interagit avec CAV1, et la cavine1 pourraient faciliter la formation de cavéoles, mais pas dans tous les tissus.
2. La composition lipidique de nanodomaines membranaires, en particulier, cholestérol et en phospholipides chargés négativement, i.e. phosphatidylsérine (PS) et PI(4,5)P2 ou PIP2, est indispensable à la biogenèse des cavéoles.
L'assemblage ainsi que les interactions entre toutes ces molécules, i.e. protéines/lipides, protéines/protéines et lipides/lipides sont loin d'être connus, ce qui implique que certains articles un peu anciens ne sont plus d'actualité et que les modèles sont nombreux.
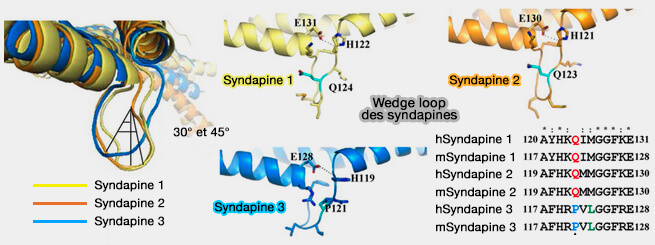
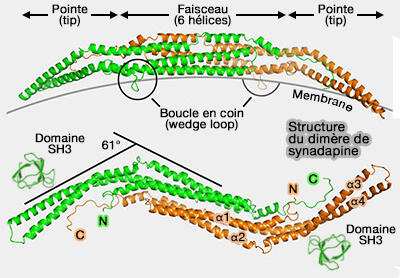
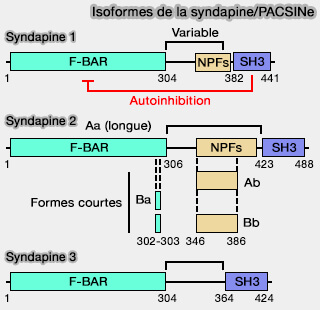
Syndapine 2/PACSINe 2
Isoformes de la syndapine/PACSINe
(Figure : vetopsy.fr d'après Quan et coll)
La syndapine 2 stabiliserait le col cavéolaire à la membrane plasmique, i.e. les cavéoles deviennent plus dynamiques et leur durée d'attachement à la membrane plasmique diminue au fur et à mesure de l'épuisement de la syndapine 2.
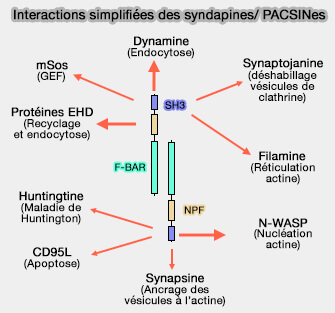
Interactions simplifiées des
syndapines/PACSINes
(Figure : vetopsy.fr d'après Wang et coll)
La dissociation directe du col des cavéoles diminue la stabilité des cavéoles au niveau de la membrane plasmique.
EHD2 et la syndapine 2, par leurs interactions, peuvent stabiliser ensemble le cou des cavéoles.
cette phosphorylation pourrait aussi recruter la dynamine pour la fission.
2. Cav1 s'associe à la filamine A qui stabilise le mouvement latéral des cavéoles, mais qui peut aussi se lier à PKCα et contribuer au détachement des syndapines ( phosphorylation de Cav1 et endocytose).
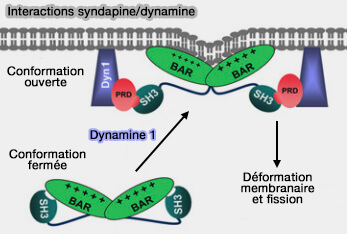
Interaction syndapine/dynamine
(Figure : vetopsy.fr d'après Rao et coll)
Ce recrutement pourrait être causé par la phosphorylation de la syndapine 2 et sa dissociation des cavéoles (et de ses interactions avec EHD2) pour recruter la dynamine.
Les cavéoles étant des microdomaines stables au niveau de la membrane plasmique, le rôle réel de la dynamine et ses interactions avec les protéines stabilisatrices du col, EHD2 et syndapine 2 doivent encore être clarifiées.
Soit ROR1 servirait de protéine d'échafaudage en se liant directement à la cavine 1 et à la cavéoline-1 (CAV1), fonction indépendante de son rôle de kinase, pour faciliter la formation de cavéoles.
Soit, il emploierait une autre protéine inconnue à ce jour.
ROR1 et formation des cavéoles
(Figure : vetopsy.fr d'après Yamagushi et coll)





 oligomérisation des syndapines),
oligomérisation des syndapines),