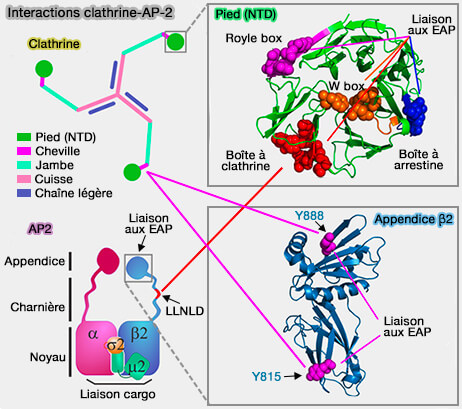
Les adaptines (AP ou Adaptor Protein) sont une famille de complexes hétérotétramères apparentés qui fonctionnent comme des revêtements pour le tri des cargos de protéines membranaires intégrales dans le système endomembranaire.
Réseau d'interactions cargo/adaptateurs
(Figure : vetopsy.fr d'après Traub et Bonifacino)
Elles engagent et accumulent des cargos sur une membrane source en se liant aux signaux de tri situés dans les domaines cytoplasmiques du cargo.
Elles recrutent une couche externe et/ou d'autres protéines accessoires pour générer des transporteurs qui facilitent la livraison du cargo à sa destination.
Les adaptines reconnaissent le récepteur par une séquence particulière de son domaine cytosolique qui est appelée peptide signal, par lequel elles se lient à lui.
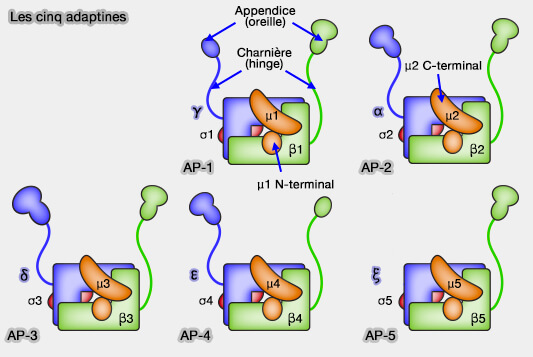
1. Ces adaptines sont des hétérotétramères (α, β, μ et σ), légèrement différents suivant les AP.
Bien qu'il y ait moins d'isoformes de sous-unités AP-2 par rapport aux sous-unités AP-1, une diversité supplémentaire provient de l'épissage alternatif de certains ARNm.
Le recrutement de la clathrine n'est pas clair, car la sous-unité β3 n'est pas concentrée dans les vésicules de clathrine du cerveau ou du foie (Forty Years of Clathrin-coated Vesicles 2015).
3. δ, β3A, μ3A, ς3A et ς3B sont exprimés de manière ubiquitaire, tandis que β3B et μ3B sont spécifiquement exprimés dans les neurones et les cellules neuroendocrines.
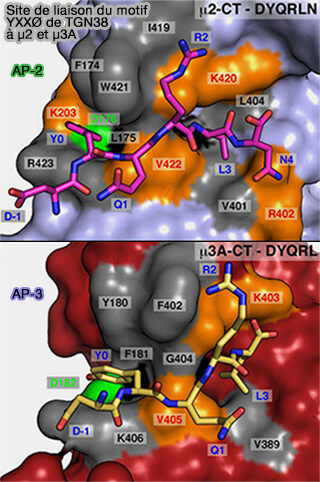
Liaison de YXXØ à μ2 et μ3A
(Figure : vetopsy.fr d'après Mardones et coll)
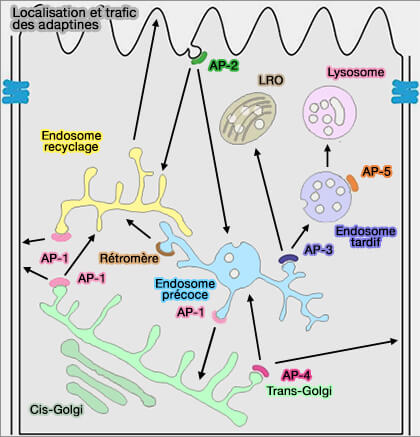
la médiation du TGN dans le trafic endosomal des cargos spécifiques, telles que la protéine précurseur amyloïde APP,
dans le tri basolatéral dans les cellules polarisées, comme celui du récepteur LDL normalement ciblé sur les dendrites somatiques, des récepteurs AMPA et δ2 du glutamate axonaux,
La voie AP-4 est donc relativement mineure par rapport à celles de AP-1 et AP-3 dans le réseau trans-Golgi (TGN).
Cependant, elle participe au trafic d'un ensemble spécifique de protéines qui représentent collectivement une fraction mineure de toute la membrane cargaisons qui sortent du TGN.
Curieusement, le déficit en AP-5 est associé à une paraplégie spastique progressive, ce qui suggère que l'AP-5, comme l'AP-4, joue un rôle fondamental dans le développement neuronal et l'homéostasie.
Les parallèles phénotypiques inattendus entre les patients AP-4 et AP-5 peuvent à leur tour suggérer une relation fonctionnelle des deux AP dans le trafic des vésicules.
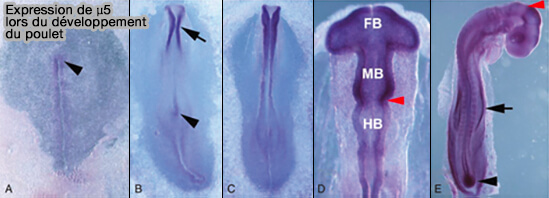
Expression de μ5 lors du développement du poulet
(Figure : vetopsy.fr d'après Hirst et coll)
b. La sortiline pourrait agir comme un lien entre les protéines du Golgi et le complexe AP-5/SPG11/SPG15, ce qui expliquerait, lors de mutations, l'accumulation d'endo-lysosomes aberrants dans les cellules et le rôle du dysfonctionnement des endosomes/lysosomes dans la pathologie de la paraplégie spastique héréditaire et d'autres troubles neurodégénératifs.




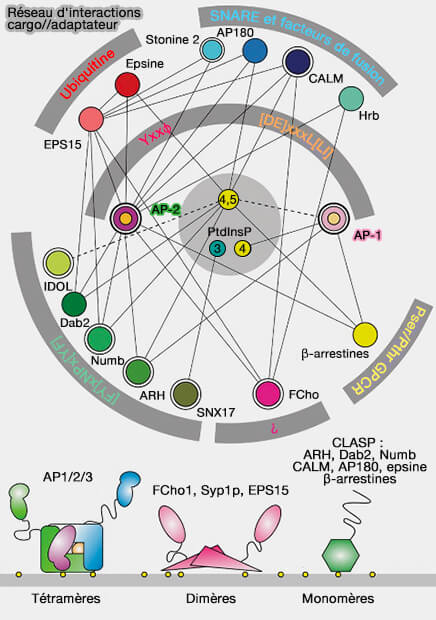

 adaptateurs alternatifs).
adaptateurs alternatifs).