L'inactivation de la formation du complexe SNARE par les protéines SNARE elles-mêmes est liée à deux domaines N-terminaux, i.e. le domaine Habc des Q-SNARE et le domaine longin des R-SNARE.
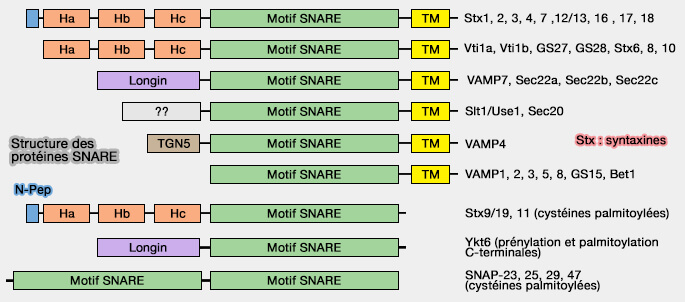
Structure des protéines SNARE
(Figure : vetopsy.fr d'après Hong et Lev)
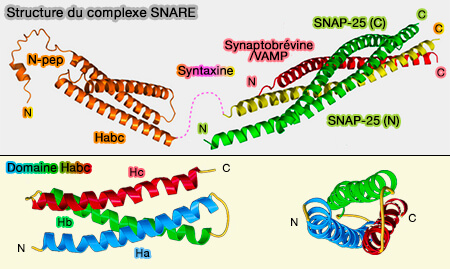
Domaine Habc
Structure
Le domaine Habc, retrouvé dans les Qa-, Qb- et certains
Qc-SNARE, forme un faisceau replié de trois hélices α (a, b et c) superposables, connecté au domaine SNARE par un long lien flexible (linker).
Structure du complexe SNARE et domaine Habc de Stx1
(Figure : vetopsy.fr d'après Stepien et coll)
Dans certaines syntaxines (Stx), en particulier dans Stx1, mais pas toutes, les domaines Habc ont la capacité de s'engager dans une interaction intramoléculaire avec le domaine SNARE pour inhiber la formation du faisceau hélicoïdal tétramérique du complexe SNARE et la fusion membranaire.
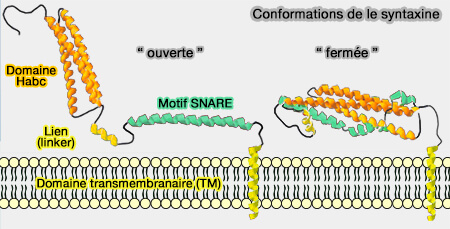
Conformations de la syntaxine
(Figure : vetopsy.fr)
1. Lorsqu'elle n'est pas liée au complexe SNARE, la syntaxine s'auto-inhibe par le repliement du domaine Habc pour masquer son motif SNARE C-terminal.
Cette conformation " fermée " inhibe la formation de complexes avec les autres protéines SNARE apparentées et donc la fusion membranaire.
Stx1 s'associe sur toute la longueur du domaine cytoplasmique de Stx1 avec Munc18, ce qui provoque la " fermeture " de Stx1 (complexe Sec1/Munc18-Qa).
On ne sait pas si la syntaxine s'auto-inhibe toute seule ou si elle nécessite Munc18 pour le faire.
Modèle d’ouverture de la syntaxine 1 en complexe avec Munc18
(Figure : vetopsy.fr d'après Stepien et coll)
3. Les domaines Habc de diverses syntaxines se sont avérés être des sites de liaison aux protéines qui régulent la fonction SNARE, y compris celles des familles Munc18, Munc13 et synaptotagmine.
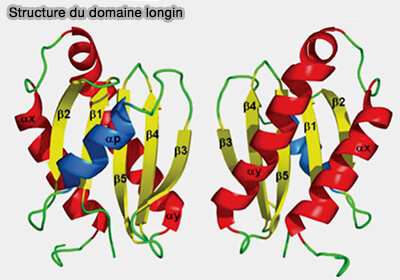
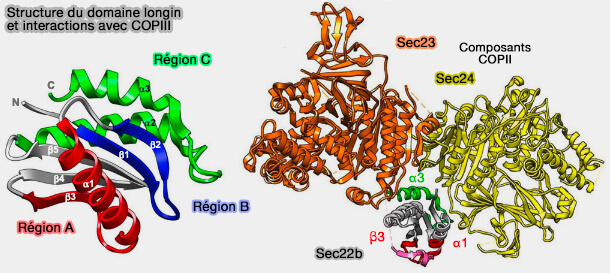
Domaine longin
Le domaine longin a d'abord été identifié comme un module de régulation des protéines de fusion membranaire des SNARE, avant d'être détecté dans six autres familles importantes de protéines de trafic.
Le domaine longin forme un pli globulaire de 120 à 140 acides résidus, disposé en sandwich α-β-α, l'un des plis les plus courants trouvés dans la nature.
2. Trois régions de liaison (régions A, B et C) ont été identifiées au sein des protéines à domaine longin.
a. La région A est impliquée dans les interactions intra- et intermoléculaires, i.e. αp, β3 et la partie N-terminale de β4 et, dans certains cas, β1 N-terminal, β5 C-terminal et αy N-terminal.
Cette région est cruciale pour réguler la fusion membranaire et la localisation subcellulaire.
b. La région B est impliquée dans les interactions avec les petites GTPases, i.e. αp et l'épingle à cheveux β1-β2 ainsi que les hélices αx-αy.
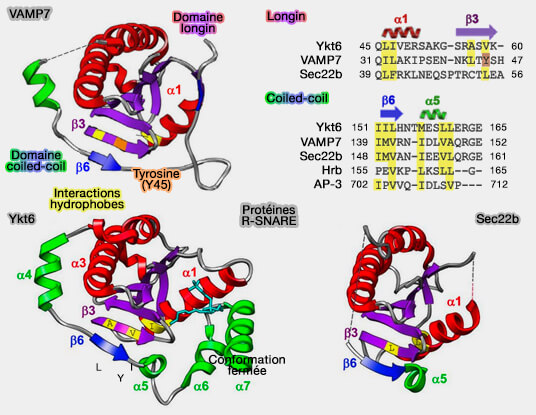
Comparaison des structures " fermées " des R-SNARE
(Figure : vetopsy.fr d'après Daste et coll)
c. La région C, impliquée dans des interactions conformationnelles avec des complexes protéiques volumineux, est la région la plus variable en raison du nombre différent d'hélices α supplémentaires.
Dans différentes superfamilles homologues de longin, cette région de liaison peut impliquer soit une hélice αx ou αy individuelle, soit les deux.
Dans les profilines, protéine qui interagit avec l'actine et dont le rôle est complexe ( profiline et actine), elle est centrée entre les deux hélices.
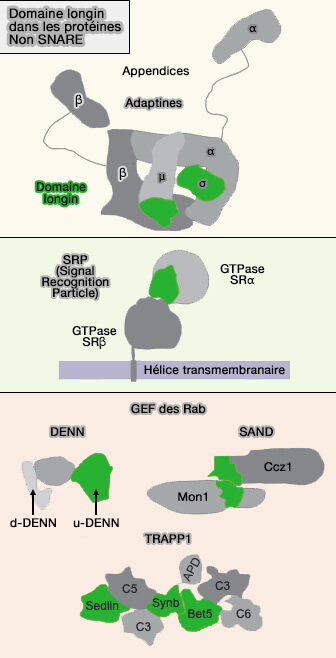
Fonctions du domaine longin
Dans les protéines non SNARE
Adaptines et COPI/II
1. Les complexes d'enveloppe vésiculaire contenant le domaine longin interviennent dans le bourgeonnement des vésicules et la sélection des cargos dans :
Dans les sédlines, le domaine longin est un composant des complexes TRAPP I et II (TRAnsport Protein Particle) qui se localisent à la surface du cis-Golgi et du trans-Golgi (TGN), respectivement, et sont nécessaires pour attacher les vésicules dérivées du réticulum endoplasmique (RE) à la membrane de Golgi et pour le trafic intra-Golgi.
Dans les complexes TRAPP I multimériques, le domaine longin est présent dans trois sous-unités, i.e. sedline/TRAPPC2, synbindine/TRAPPC4 (Synb) et Bet5/TRAPPC1.
Les structures cristallines de ces sous-unités révèlent un domaine longin avec une forte similitude avec celui des SNARE, malgré l'absence de similitude de séquence.
Targetines
Dans les targetines, le domaine longin est exprimé de manière différentielle dans les cellules normales et néoplasiques (DENN) et les protéines létales synthétiques APL2 VPS1 (AVL).
Ces GEF (Guanine nucleotide exchange factor) sont importantes chez les eucaryotes supérieurs, dans lesquels le domaine longin régulateur N-terminal (domaine u-DENN) est adjacent au domaine DENN du facteur d'échange GTP-GDP.
Le module DENN tripartite, i.e. domaine longin N-terminal, domaines DENN et d-DENN, est un GEF pour les petites GTPases Rab, qui sont des régulateurs de pratiquement tous les événements de trafic membranaire chez les eucaryotes.
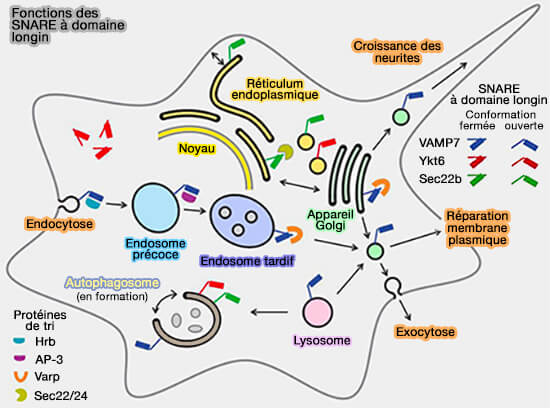
Dans les protéines SNARE
Le domaine longin est retrouvé dans trois familles de protéines Q-SNARE qui sont traitées dans des chapitres spéciaux, i.e. VAMP7, Sec22 et Ykt6.
Fonctions des SNARE à domaine longin
(Figure : vetopsy.fr d'après Daste et coll)
dans les interactions intramoléculaires avec le domaine SNARE (pour réguler son activité),
dans la liaison intermoléculaire à d'autres protéines de trafic afin de cibler les longin des SNARE vers leur site d'action.
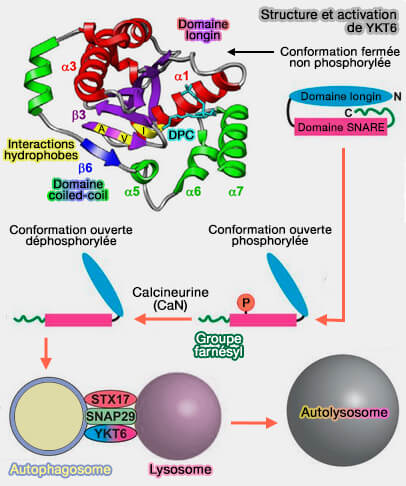
Ykt6 et son activation
(Figure : vetopsy.fr d'après McGath et coll)
2. VAMP7 et Ykt6, seuls, peuvent adopter une conformation fermée et stable qui altère l'assemblage du complexe SNARE.
Vamp7 s'auto-inhibe par l'interaction de son domaine longin avec son propre domaine SNARE, renforcé par Varp ( activation de VAMP7).
Cette conformation " fermée " majoritaire en solution lui permet son déplacement au sein de la cellule.
La phosphorylation de la tyrosine 45 (Y45) de β3 provoque la transition de sa position fermée à la position ouverte.
Ykt6 s'auto-inhibe aussi par l'interaction de son domaine longin avec son propre domaine SNARE ( activation de Ykt6).
Cette conformation inactive de Ykt6 est stabilisée par le groupe farnésyl, pris en sandwich entre le domaine SNARE et le domaine longin.
La phosphorylation sur un site conservé au cours de l'évolution dans le domaine SNARE peut réguler un changement conformationnel d'une forme cytosolique fermée à une forme plus ouverte liée à la membrane.




 complexe Sec1/Munc18-Qa).
complexe Sec1/Munc18-Qa)..jpg)