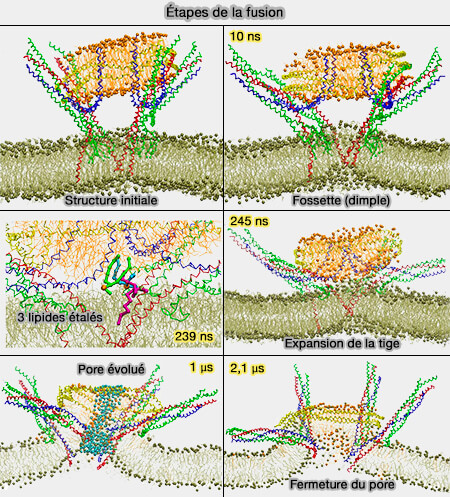
Les protéines de fusion sont censées déformer une petite zone de membrane pour former des structures en forme de pointes, i.e. la tige de fusion, qui font saillie vers le partenaire de fusion et favorisent l'étalement des lipides des feuillets externes.
2. L'hémifusion est l'étape dans laquelle les feuillets externes sont fusionnés.
Les lipides peuvent passer entre les feuillets externes, mais les feuillets internes séparés empêchent le mélange des contenus.
Toutefois, l'hémifusion pourrait être en équilibre avec de petits pores de fusion qui oscillent ( évolution du pore de fusion).
L'hémifusion semble être un intermédiaire universel qui peut être observé dans les réactions de fusion entre bicouches purement lipidiques ainsi que dans la fusion protéique entre membranes physiologiques (Mechanics of membrane fusion 2008).
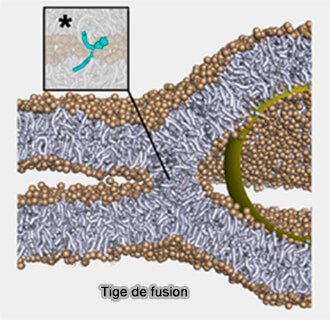
Tige de fusion (hémifusion)
(Figure : vetopsy.fr d'après Grubmüller et coll)
Cette expansion ne se déroule pas nécessairement de manière radiale, on peut aussi utiliser le terme d'élargissement de la tige (stalk widening) qui est plus approprié.
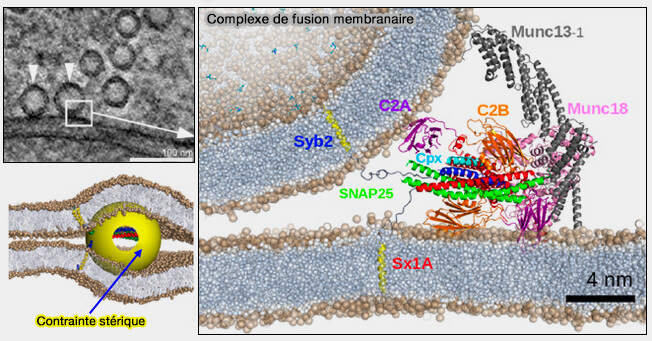
La présence supplémentaire d'un seul complexe d'attache volumineux comme le complexe HOPS associé aux SNARE peut déjà réduire la barrière d'énergie libre contre l'élargissement de la tige à environ 35 kBT en imposant des répulsions stériques sur le site de l'hémifusion.
Ces complexes d'attache imposent un emplacement périphérique du complexe SNARE près du bord incurvé de la membrane (le sommet) de la zone de contact/adhésion formée entre les grandes vésicules.
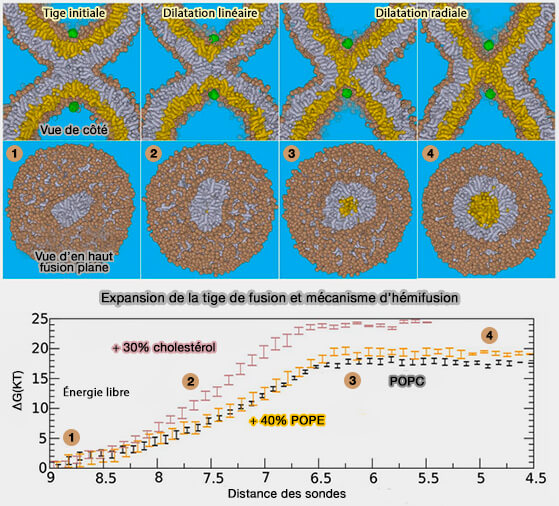
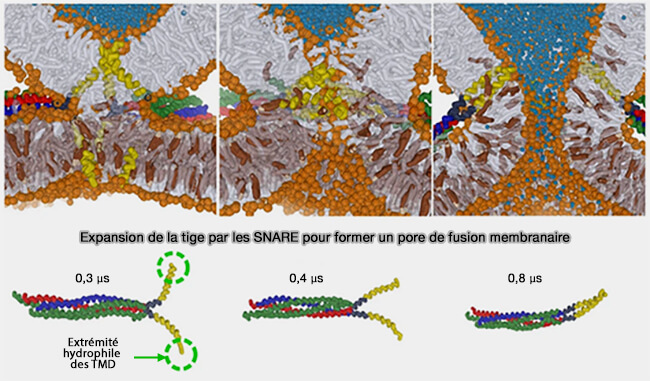
b. La figure ci-dessous montre la réaction de fusion entre les membranes en réponse à la traction de deux sondes hydrophiles (vert) l'une vers l'autre à travers le centre de la tige, i. e. ce processus imite l'action de la fermeture éclair SNARE où les extrémités C-terminales du domaine transmembranaire hydrophile sont rapprochées.
La tige initiale (1) se dilate linéairement (2) avant de se dilater radialement (3 et 4). Une fois la barrière (3) de 17 à 24 kBT franchie, la formation/l'expansion ultérieure du diaphragme d'hémifusion (4) devient spontanée, i.e. région du plateau.
L'énergie libre requise est représentée en noir sur le bas de la figure, en jaune avec l'ajout de POPE (1-palmitoyl-2-oléoyl-phosphatidyléthanolamine), à effet relativement neutre, et de cholestérol qui augmente marginalement la barrière contre l'élargissement de la tige, de 17 à 24 kBT ( lipides membranaires et tige de fusion).
Les feuillets internes sont représentés en jaune, les feuillets externes en gris, et l'exemple montre la situation pour la membrane POPC (1-palmitoyl-2-oléoyl-phosphatidylcholine) pure, i.e. les groupes de tête PC sont représentés en brun.
Expansion de la tige de fusion et mécanisme d’hémifusion
(Figure : vetopsy.fr d'après Risselada et coll)
2. Les barrières contre l'élargissement de la tige peuvent être surmontées par :
Ce n'est pas seulement la barrière contre l'expansion de la tige qui constitue un défi dans la fusion synaptique, c'est plutôt l'exigence d'une réaction de fusion rapide, i.e. en quelques millisecondes et exclusivement sans fuite.
3. La machinerie de fusion peut appliquer activement une telle réaction de deux manières.
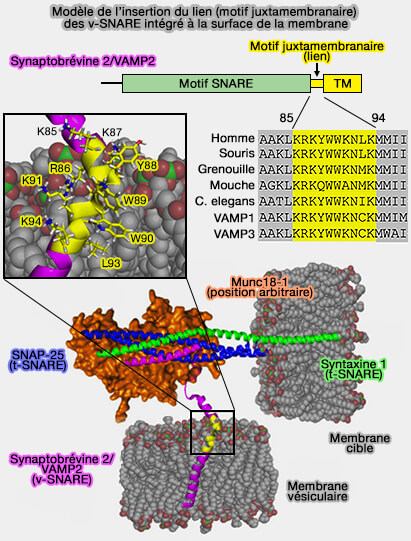
a. Soit, elle peut imposer une proximité étroite du feuillet interne en soumettant une force ponctuelle sur le feuillet, par exemple, via les domaines transmembranaires (TMD) des SNARE.
b. Soit, elle peut activement condenser les feuillets externes près du site de fusion, par exemple via la condensation de lipides chargés négativement en présence de Ca++.
La présence des liens des protéines SNARE chargés positivement, qui se localisent près des groupes de tête lipidiques, peut en outre contribuer à une telle condensation électrostatique.
La machinerie de fusion synaptique rapide semble impliquer une condensation (locale) des feuillets externes pour imposer l'élargissement de la tige.
3. L'élargissement de la tige lors de la fusion virale est spécial, car les protéines de fusion virales sont apparemment incapables d'exercer une force mécanique directe sur les deux feuillets internes de la tige, car cela nécessiterait que leurs peptides de fusion amphiphiles pénètrent d'abord et traversent complètement la membrane de l'hôte.
La tige peut être élargie via la formation d'un " trou " dans la membrane cible à proximité directe de la tige, ce qui provoque la réduction mutuelle de l'énergie libre de la tige et du " trou ".
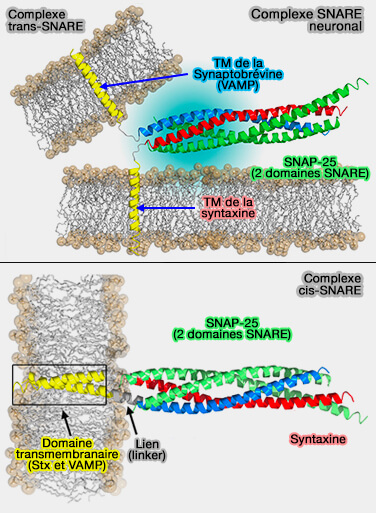
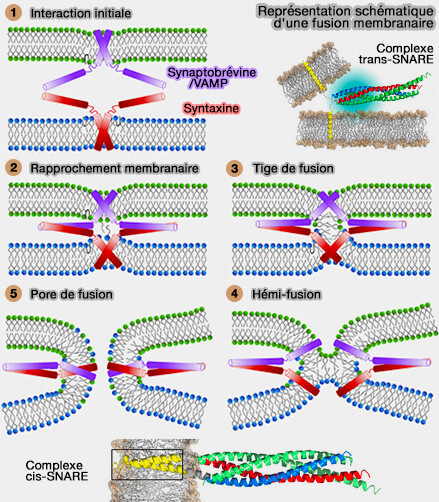
Complexe SNARE neuronal
(Figure : vetopsy.fr d'après Fang et Lindau)
En raison de la tension excessive de la ligne présente au bord du " trou ", la tige l'encercle rapidement en formant un diaphragme d'hémifusion.
Les complexes SNARE peuvent activement surmonter la barrière contre l'élargissement de la tige en imposant mécaniquement une pression locale sur les feuillets internes via les extrémités hydrophiles des domaines transmembranaires (TMD), ce qui facilite leur indentation locale.
Ce mécanisme repose essentiellement sur un équilibre de force élastique entre la force générale associée à la croissance de la surface de la tige et la force ponctuelle imposée par les extrémités des TMD sur les feuillets internes.
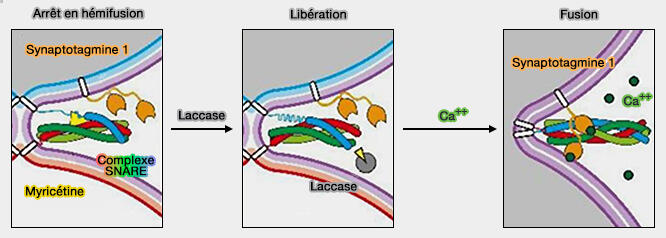
La formation du complexe SNARE peut être arrêtée à l'état semi-zippé par la myricétine, un flavonoïde anti-oxydant, i.e. cet état correspond à l'hémifusion dans la fusion du liposome.
La myricétine bloque la fermeture à glissière C-terminale en se liant à la région médiane, tout en permettant la fermeture des régions N-terminales des SNARE.
Même lorsque les membranes sont à de petites distances de séparation intermembranaires, la barrière d'énergie libre pour former la structure de la tige est comprise entre 17 et 24 kBT.
Les TMD des SNARE peuvent induire une réduction substantielle (∼10 kBT) de l'énergie libre de la formation de la tige et de la barrière contrôlant sa transition vers un pore de fusion.
Cela est dû à l'amincissement local de la membrane, fossette pré-tige, imposé par les TMD à proximité de la structure de la tige fortement courbée.
Cet effet peut être relativement indépendant de la séquence, mais lié à la longueur hydrophobe effective du TMD, i.e. le TDM peut exprimer un décalage hydrophobe avec la membrane, de sorte que sa longueur est mieux adaptée dans un pédoncule que dans une simple bicouche.
2. La distance entre les membranes est le déterminant le plus crucial de l'énergie libre de la tige.
La forme des lipides joue un rôle crucial pour surmonter la répulsion d'hydratation entre deux membranes et diminue le travail total requis pour former une tige.





 machinerie de fusion).
machinerie de fusion).