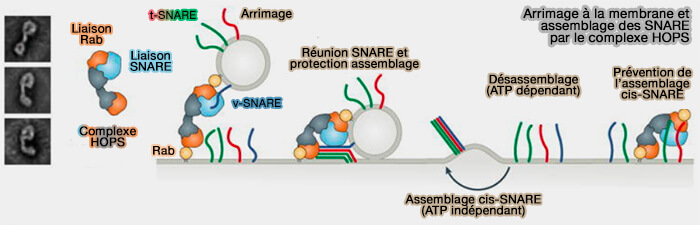
Le rapprochement des membranes dans la fusion membranaire est lié aux protéines SNARE, mais aussi aux Rab, aux protéines d'amarrage (tethering) et aux protéines SM.
3. Le complexe d'amarrage reconnait l'autre membrane via un deuxième site de liaison, éventuellement une autre Rab ou des phosphoinositides spécifiques, et attache les deux membranes.
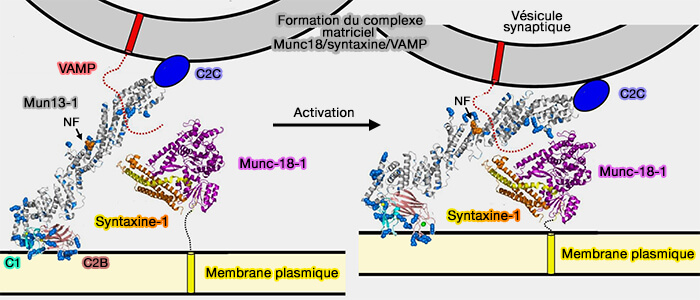
Attache à la membrane et assemblage des SNARE par le complexe HOPS
(Figure : vetopsy.fr d'après Baker et Hugson)
3. Ce complexe possède plusieurs sites de liaison SNARE et favorise l'assemblage local des protéines SNARE en faisceaux à quatre hélices, en stabilisant leurs conformations ouvertes et en chaperonnant l'assemblage via les protéines Sec1/Munc18-like (SM) associées.
Les complexes d'amarrage relient la machinerie de régulation Rab à la machinerie de fusion SNARE, mais les interactions sont difficiles à étudier car elles évoluent pendant le déroulement de la fusion membranaire (rôle des complexes d'attache multi-sous-unités dans l'assemblage SNARE).
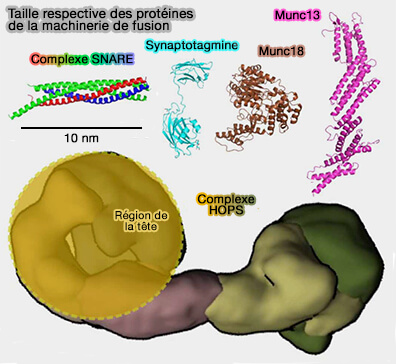
Taille des protéines de fusion
(Figure : vetopsy.fr d'après Risselada et Mayer)
l'accessibilité des SNARE aux membranes qui les entourent,
leur orientation et leur conformation,
la topologie des membranes qui les entourent,
les changements conformationnels et les forces qu'il peut imposer sur les deux membranes à fusionner.
Les SNARE sont relativement petites, i.e. inférieures à 100 kDa assemblées, alors que les protéines associées sont volumineuses, comme les SM (100 kDa) ou les MTC (0,25–1 MDa).
2. L'étape principale de la fusion membranaire doit être précédée d'une approche des membranes opposées en dessous d'une distance critique d'env. 1 nm, quelle que soit la composition de la membrane ou la contrainte stérique imposée (Vesicle Adhesion and Fusion Studied by Small-Angle X-Ray Scattering 2018).
a. Or, les complexes protéiques volumineux liés aux SNARE modifient la distance et la courbure des membranes au site de fusion et ont ainsi un impact significatif sur le paysage énergétique de la formation des pores de fusion et de leur expansion.
Un défi stérique encore plus important est posé par la fusion endosome-vacuole tardive, où la courbure impliquée au site de fusion membranaire est beaucoup plus faible et les complexes liés à SNARE, i.e. le complexe HOPS, sont encore plus grands.
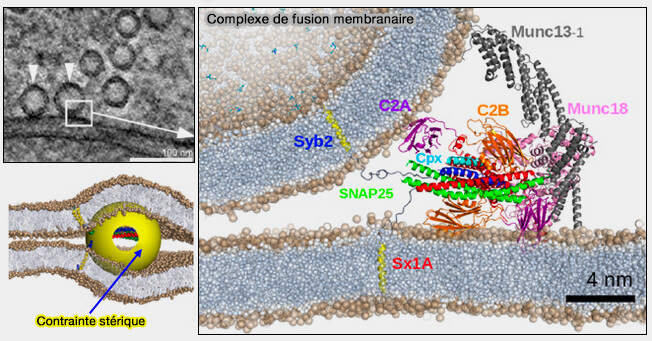
Complexe de fusion membranaire
(Figure : vetopsy.fr d'après Grubmüller et coll)
2. Les complexes d'attache interagissent avec les membranes par le biais d'interactions protéine-protéine et par une affinité directe pour les lipides membranaires.
a. Bien que ces interactions puissent maintenir les membranes à proximité les unes des autres, il n'y a aucune preuve que les protéines d'attache déforment activement les membranes au niveau d'un site de fusion.
b. Par contre, les protéines SNARE courbent les membranes pour apposer les deux membranes.
Cela augmente le travail nécessaire quand elles sont attachées à un gros complexe protéique pour former la tige de fusion.
en positionnant les SNARE de telle sorte que la formation du complexe SNARE soit stable, comme par exemple, la protéine SM Vps33 trouvée dans complexes endo-lysosomaux…,
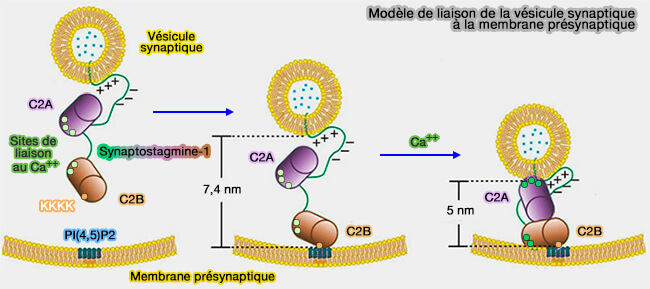
Modèle de liaison de la vésicule synaptique à la membrane présynaptique
(Figure : vetopsy.fr d'après Lin et coll)
5. L'interaction des complexes SNARE avec ces protéines influence fortement l'accessibilité des SNARE, l'orientation et la conformation des complexes SNARE, et la topologie des membranes qui les entourent.
Les protéines associées à SNARE auront un impact majeur sur la forme et les dimensions de l'holo-complexe, et sur les changements conformationnels et les forces qu'il peut imposer sur les deux membranes à fusionner ( encombrement stérique et complexe SNARE).
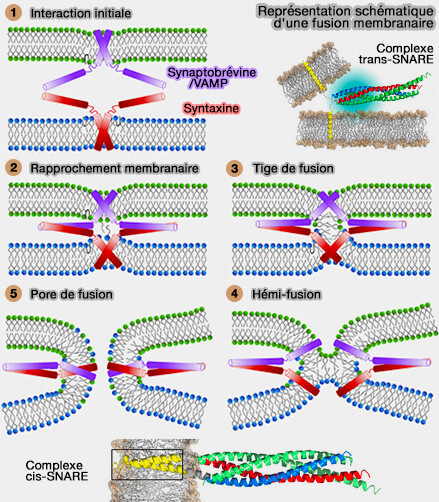
Complexes trans-SNARE
Formation du complexe trans-SNARE
1. L'interaction des protéines SNARE, les protéines v-SNARE (v pour vésiculaire) et les protéines t-SNARE (t pour target, cible), localisées sur des membranes séparées, construit un assemblage progressif en forme de fermeture éclair des quatre domaines SNARE, en général un de chaque sous-famille (Qa/Qb/Qc et R), formant le complexe trans-SNARE.
1. La déformation des membranes par les protéines SNARE a un coût énergétique.
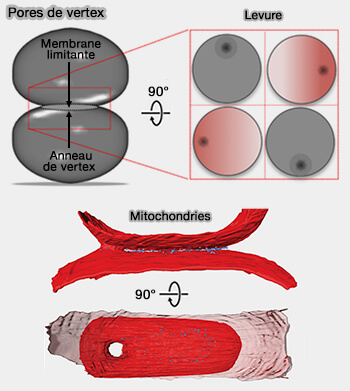
Pores de vertex
(Figure : vetopsy.fr d'après D'agostino et Brandt)
a. Pour réduire ce coût, les complexes SNARE peuvent s'accumuler au bord incurvé de la zone de contact membranaire ( pore de vertex).
b. L'association des complexes SNARE à des protéines volumineuses entraîne encore plus efficacement les complexes SNARE vers le bord de la zone de contact, dans laquelle le volume peut le plus facilement s'adapter.
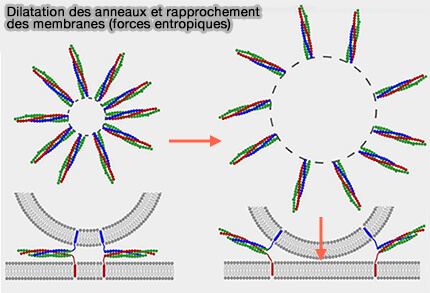
Dilatation des anneaux et rapprochement des membranes
(Figure : vetopsy.fr d'après Mostafavi et coll)
Ces interactions augmentent la distance entre les groupes et forcent les membranes à entrer en contact ( courbure membranaire et crowding).
Plus les complexes SNARE sont nombreux, i.e. par exemple, 70 v-SNARE sont disponibles par vésicule, plus les forces entropiques sont importantes, i.e. les forces entropiques sous-entendent les forces liées à l'encombrement moléculaire.
Ce mécanisme est valable aussi pour les complexes SNARE/HOPS, qui confinent les complexes SNARE associés à la région du vertex, i.e. augmentent efficacement leur concentration en étendant la zone de contact.





 machinerie de fusion).
machinerie de fusion).